

Structure of the Munc18c/Syntaxin4 N-peptide complex defines universal features of the N-peptide binding mode of Sec1/Munc18 proteins
- PMID: 17517664
- PMCID: PMC1885578
- DOI: 10.1073/pnas.0701124104
Structure of the Munc18c/Syntaxin4 N-peptide complex defines universal features of the N-peptide binding mode of Sec1/Munc18 proteins
Abstract
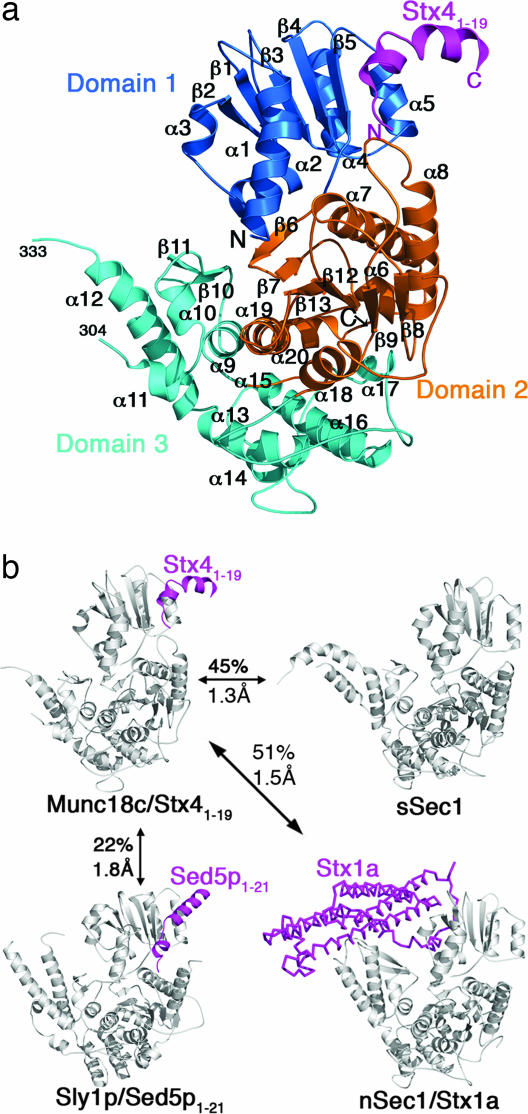
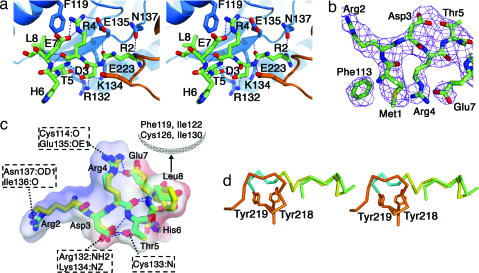
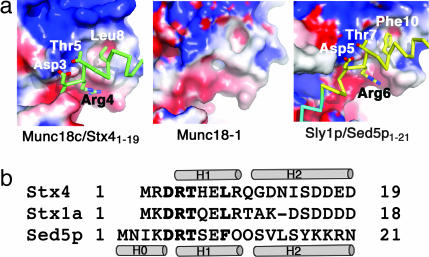
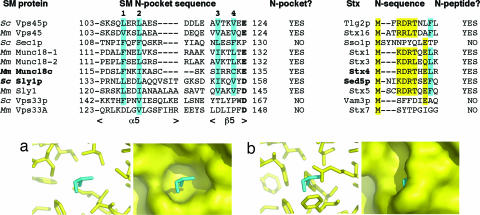
Sec1/Munc18 proteins (SM proteins) bind to soluble NSF attachment protein receptors (SNAREs) and play an essential role in membrane fusion. Divergent modes of regulation have been proposed for different SM proteins indicating that they can either promote or inhibit SNARE assembly. This is in part because of discrete modes of binding that have been described for various SM/SNARE complexes. One mode suggests that SM proteins bind only to Syntaxins (Stx) preventing SNARE assembly, whereas in another they facilitate SNARE assembly and bind to SNARE complexes. The mammalian cell surface SM protein Munc18c binds to an N-peptide in Stx4, and this is compatible with its interaction with SNARE complexes. Here we describe the crystal structure of Munc18c in complex with the Stx4 N-peptide. This structure shows remarkable similarity with a yeast complex indicating that the mode of binding, which can accommodate SNARE complexes, is highly conserved throughout evolution. Modeling reveals the presence of the N-peptide binding mode in most but not all yeast and mammalian SM/Stx pairs, suggesting that it has coevolved to fulfill a specific regulatory function. It is unlikely that the N-peptide interaction alone accounts for the specificity in SM/SNARE binding, implicating other contact surfaces in this function. Together with other data, our results support a sequential two-state model for SM/SNARE binding involving an initial interaction via the Stx N-peptide, which somehow facilitates a second, more comprehensive interaction comprising other contact surfaces in both proteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bennett MK, Garcia-Arraras JE, Elferink LA, Peterson K, Fleming AM, Hazuka CD, Scheller RH. Cell. 1993;74:863–873. - PubMed
-
- Brunger AT. Q Rev Biophys. 2005;38:1–47. - PubMed
-
- Sutton RB, Fasshauer D, Jahn R, Brunger AT. Nature. 1998;395:347–353. - PubMed
-
- Rizo J, Sudhof TC. Nat Rev Neurosci. 2002;3:641–653. - PubMed
-
- Jahn R, Scheller RH. Nat Rev Mol Cell Biol. 2006;7:631–643. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
