

Three-dimensional model for the isolated recombinant influenza virus polymerase heterotrimer
- PMID: 17517766
- PMCID: PMC1920261
- DOI: 10.1093/nar/gkm336
Three-dimensional model for the isolated recombinant influenza virus polymerase heterotrimer
Abstract
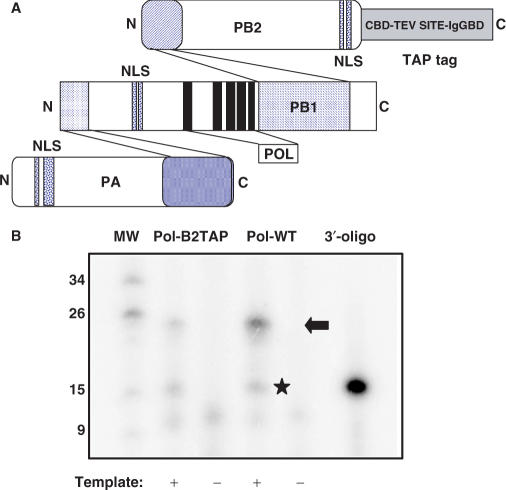
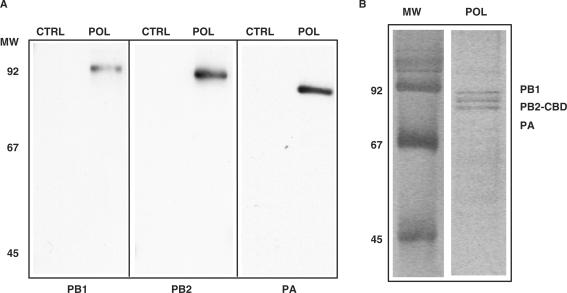
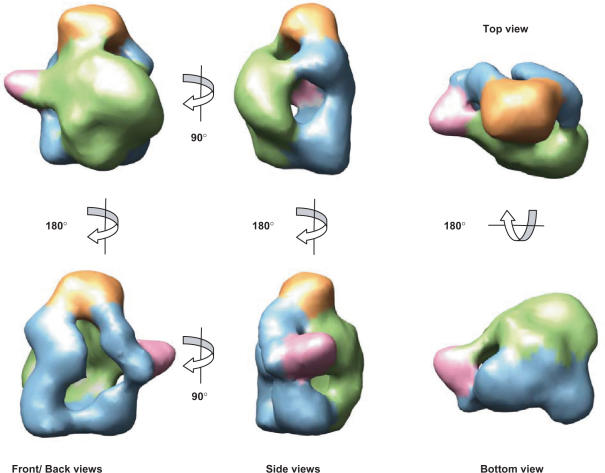
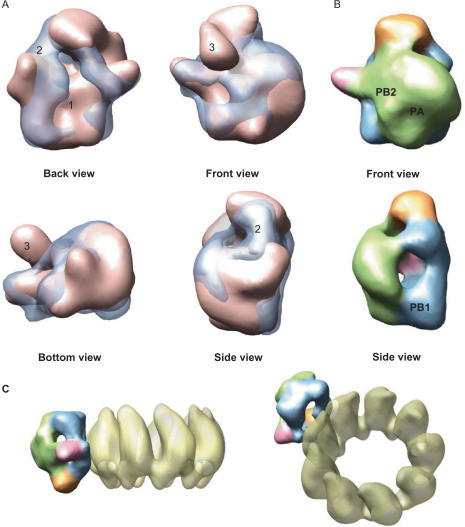
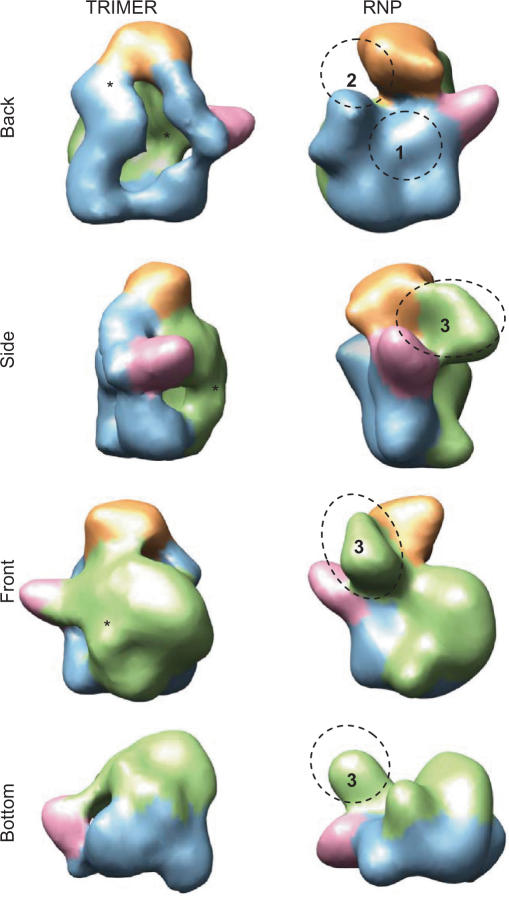
The genome of influenza A virus is organized into eight ribonucleoprotein complexes (RNPs), each containing one RNA polymerase complex. This RNA polymerase has also been found non-associated to RNPs and is possibly involved in distinct functions in the infection cycle. We have expressed the virus RNA polymerase complex by co-tranfection of the PB1, PB2 and PA genes in mammalian cells and the heterotrimer was purified by the TAP tag procedure. Its 3D structure was determined by electron microscopy and single-particle image processing. The model obtained resembles the structure previously reported for the polymerase complex associated to viral RNPs but appears to be in a more open conformation. Detailed model comparison indicated that specific areas of the complex show important conformational changes as compared to the structure for the RNP-associated polymerase, particularly in regions known to interact with the adjacent NP monomers in the RNP. Also, the PB2 subunit seems to undergo a substantial displacement as a result of the association of the polymerase to RNPs. The structural model presented suggests that a core conformation of the polymerase in solution exists but the interaction with other partners, such as proteins or RNA, will trigger distinct conformational changes to activate new functional properties.
Figures

References
-
- Palese P, Shaw M. In: Fields Virology. Howley PM, editor. 2006. Vol. 1, 5th edn. Lippincott Williams & Wilkins, Philadelphia, pp. 1647–1689.
-
- Elton D, Digard P, Tiley L, Ortín J. In: Current Topics in Influenza Virology. Kawaoka Y, editor. Norfolk: Horizon Scientific Press; 2005. pp. 1–92.
-
- Krug RM, Broni BA, Bouloy M. Are the 5’-ends of influenza viral mRNAs synthesized in vivo donated by host mRNAs? Cell. 1979;18:329–334. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
