

Proteolytic disassembly is a critical determinant for reovirus oncolysis
- PMID: 17519890
- PMCID: PMC7185731
- DOI: 10.1038/sj.mt.6300207
Proteolytic disassembly is a critical determinant for reovirus oncolysis
Abstract
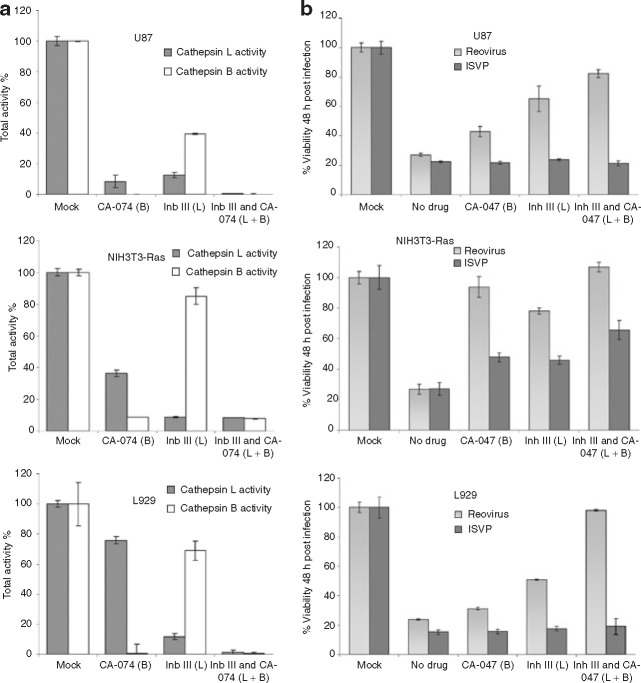
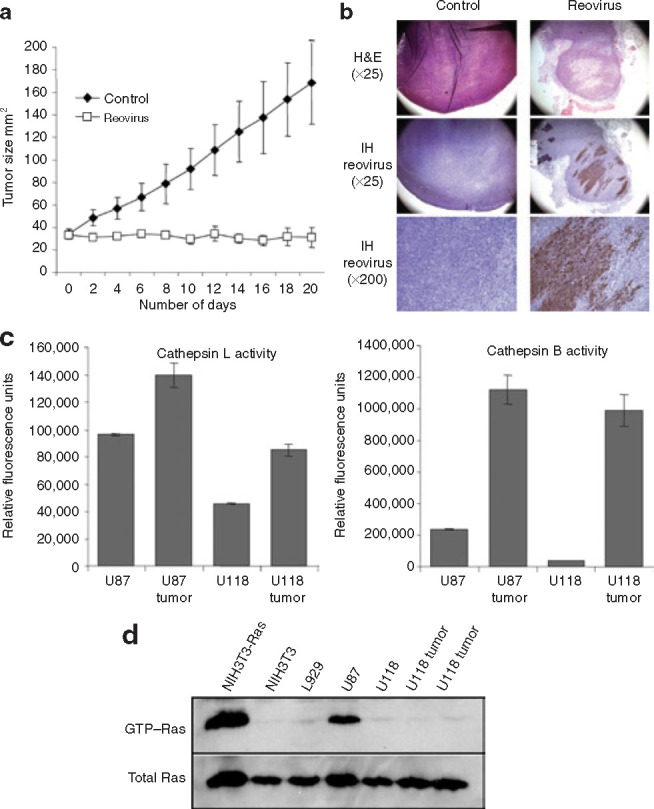
Mammalian ortheoreoviruses are currently being investigated as novel cancer therapeutics, but the cellular mechanisms that regulate susceptibility to reovirus oncolysis remain poorly understood. In this study, we present evidence that virion disassembly is a key determinant of reovirus oncolysis. To penetrate cell membranes and initiate infection, the outermost capsid proteins of reovirus must be proteolyzed to generate a disassembled particle called an infectious subviral particle (ISVP). In fibroblasts, this process is mediated by the endo/lysosomal proteases cathepsins B and L. We have analyzed the early events of infection in reovirus-susceptible and -resistant cells. We find that, in contrast to susceptible glioma cells and Ras-transformed NIH3T3 cells, reovirus-resistant cancer cells and untransformed NIH3T3 cells restrict virion uncoating and subsequent gene expression. Disassembly-restrictive cells support reovirus infection, as in vitro-generated ISVPs establish productive infection, and pretreatment with poly(I:C) does not prevent infection in cancer cells. We find that the level of active cathepsin B and L is increased in tumors and that disassembly-restrictive glioma cells support reovirus oncolysis when grown as a tumor in vivo. Together, these results provide a model in which proteolytic disassembly of reovirus is a critical determinant of susceptibility to reovirus oncolysis.
Figures
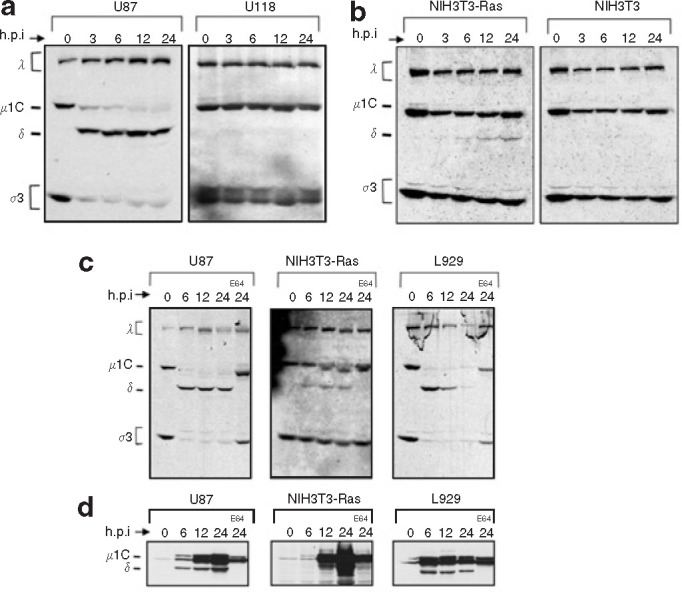
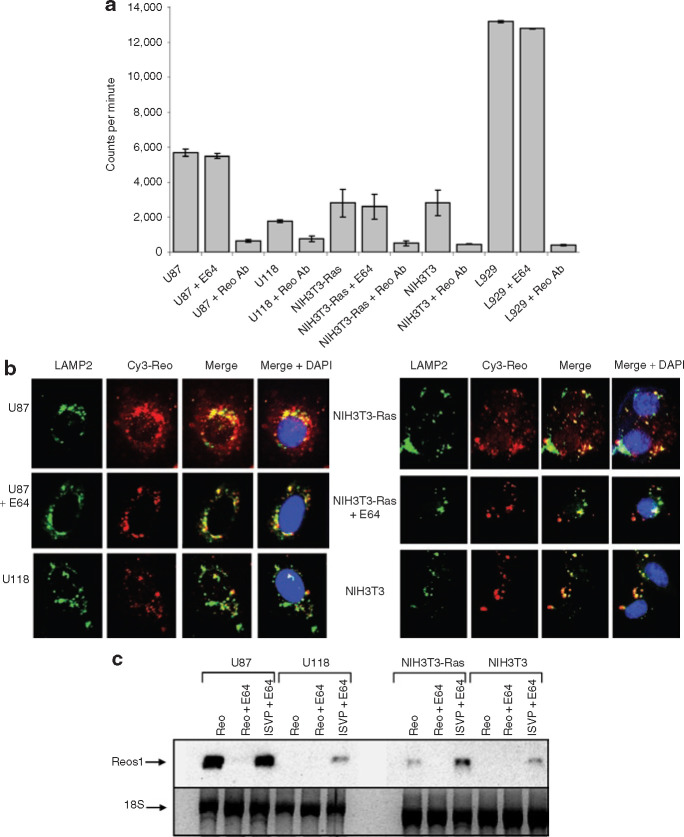
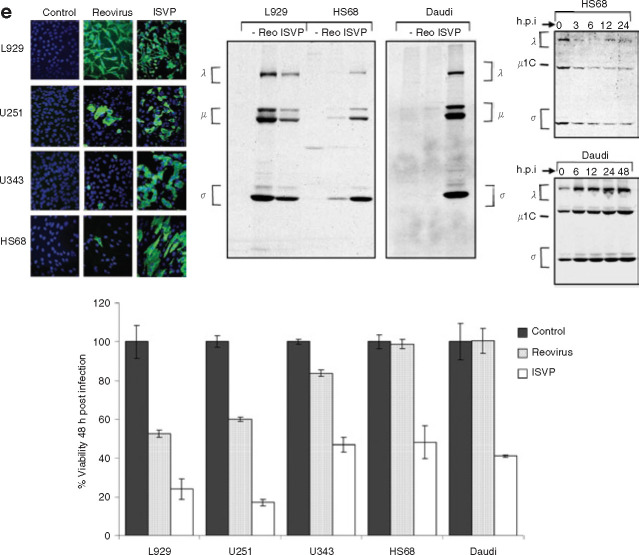
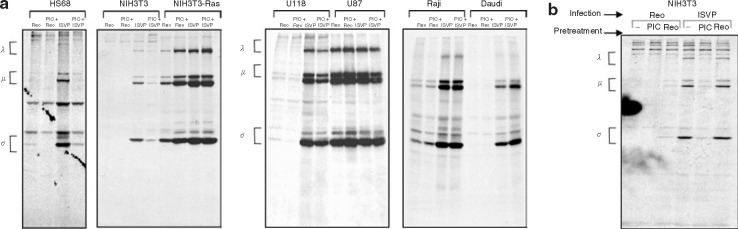
Comment in
-
Uncoating reo: uncovering the steps critical for oncolysis.Mol Ther. 2007 Aug;15(8):1406-7. doi: 10.1038/sj.mt.6300242. Mol Ther. 2007. PMID: 17646836 No abstract available.
References
-
- Tyler KL, Fields B. Reoviruses. Lippincott-Raven; Philadelphia, PA: 1996.
-
- Jackson GG, Muldoon RL. Viruses causing common respiratory infection in man. IV. Reoviruses and adenoviruses. J Infect Dis. 1973;128:811–866. - PubMed
-
- Sabin AB. Reoviruses. A new group of respiratory and enteric viruses formerly classified as ECHO type 10 is described. Science. 1959;130:1387–1389. - PubMed
-
- Rosen L, Evans HE, Spickard A. Reovirus infections in human volunteers. Am J Hyg. 1963;77:29–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
