

Plasmodium falciparum Pf34, a novel GPI-anchored rhoptry protein found in detergent-resistant microdomains
- PMID: 17521656
- PMCID: PMC2712672
- DOI: 10.1016/j.ijpara.2007.03.013
Plasmodium falciparum Pf34, a novel GPI-anchored rhoptry protein found in detergent-resistant microdomains
Abstract
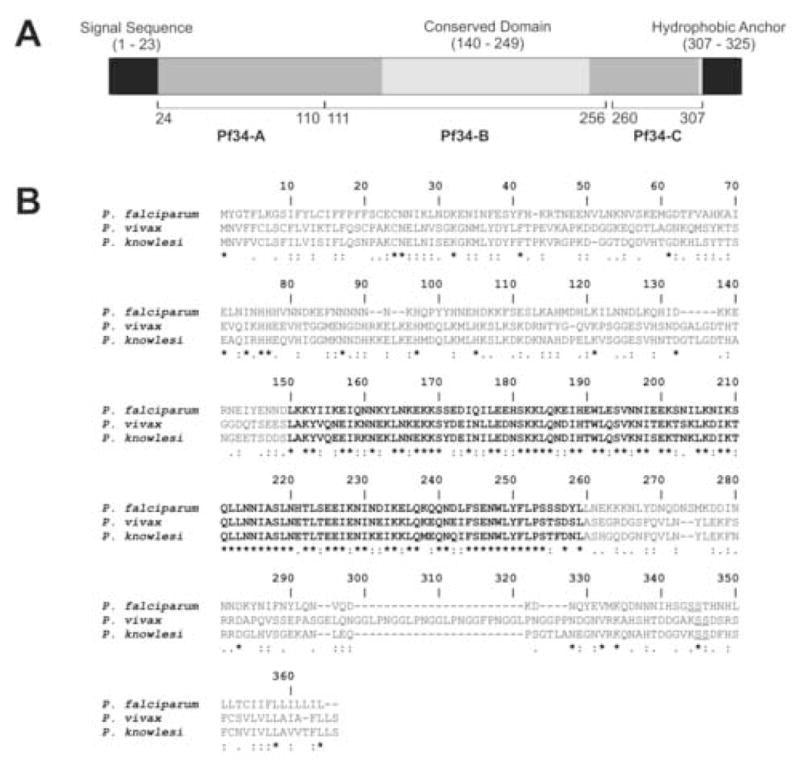
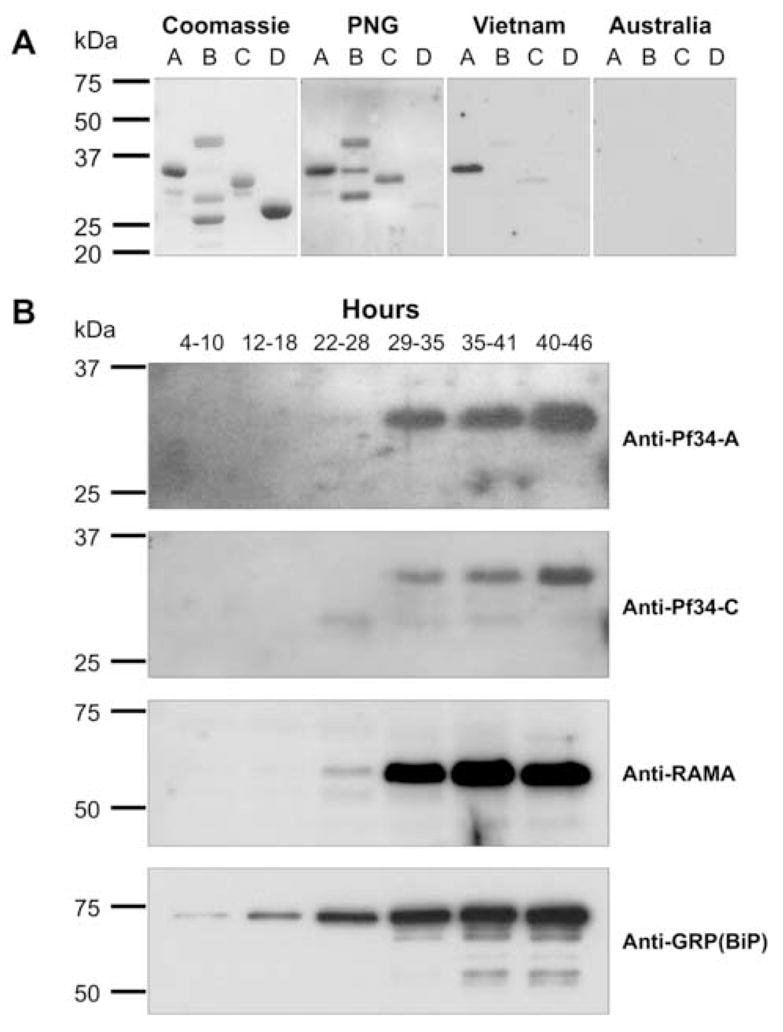
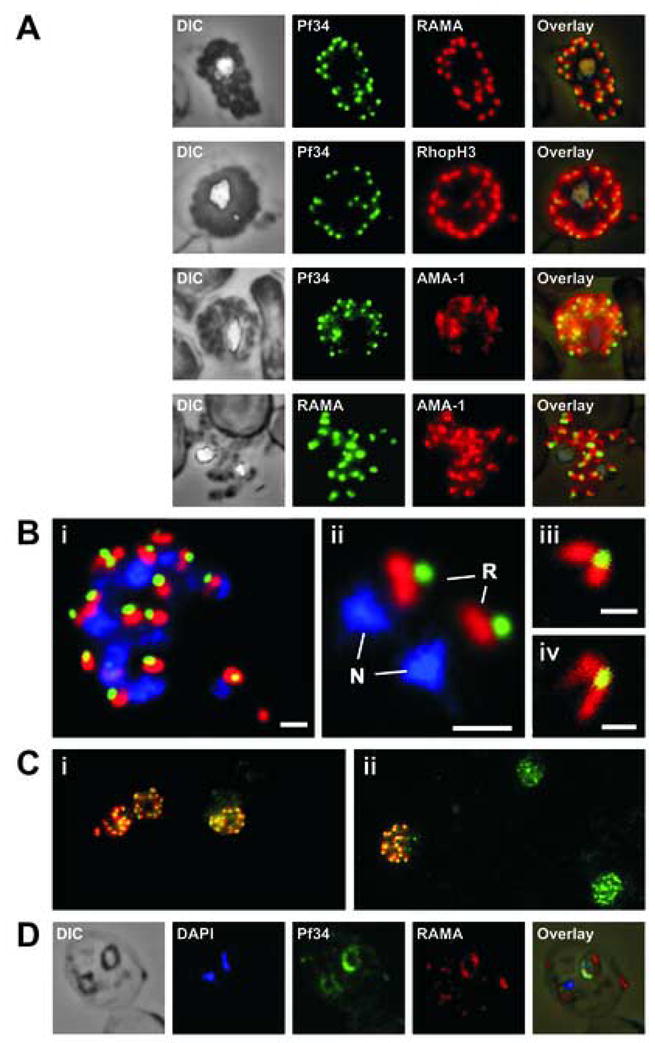
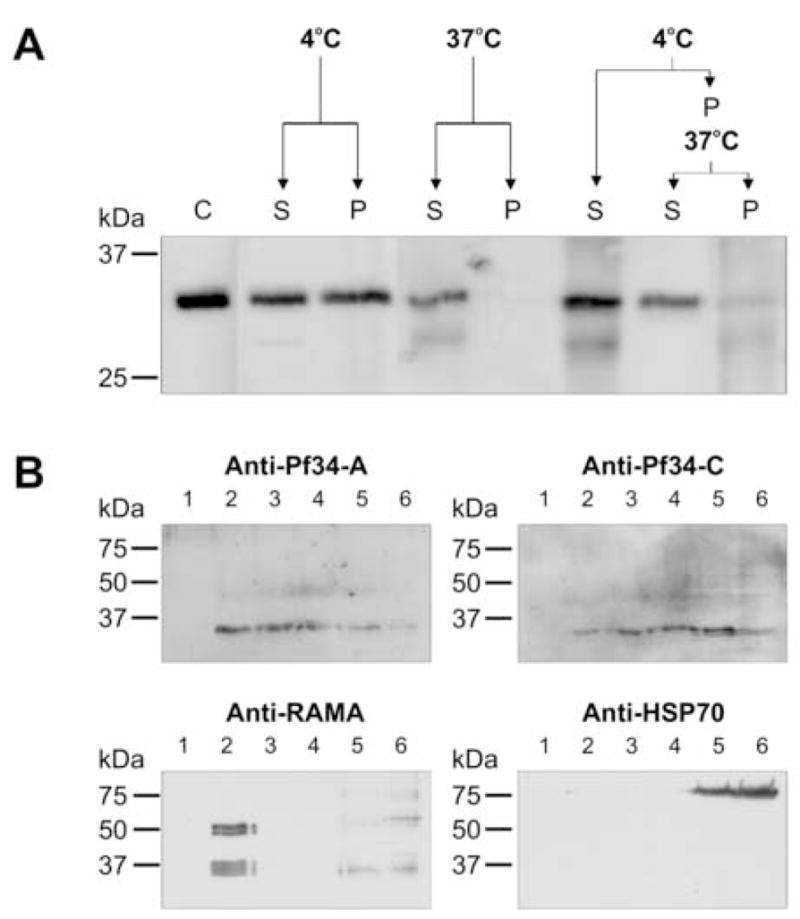
Apicomplexan parasites are characterised by the presence of specialised organelles, such as rhoptries, located at the apical end of invasive forms that play an important role in invasion of the host cell and formation of the parasitophorous vacuole. In this study, we have characterised a novel Plasmodium falciparum rhoptry protein, Pf34, encoded by a single exon gene located on chromosome 4 and expressed as a 34kDa protein in mature asexual stage parasites. Pf34 is expressed later in the life cycle than the previously described rhoptry protein, Rhoptry Associated Membrane Antigen (RAMA). Orthologues of Pf34 are present in other Plasmodium species and a potential orthologue has also been identified in Toxoplasma gondii. Indirect immunofluorescence assays show that Pf34 is located at the merozoite apex and localises to the rhoptry neck. Pf34, previously demonstrated to be glycosyl-phosphatidyl-inositol (GPI)-anchored [Gilson, P.R., Nebl, T., Vukcevic, D., Moritz, R.L., Sargeant, T., Speed, T.P., Schofield, L., Crabb, B.S. (2006) Identification and stoichiometry of GPI-anchored membrane proteins of the human malaria parasite Plasmodium falciparum. Mol. Cell. Proteomics 5, 1286-1299.], is associated with parasite-derived detergent-resistant microdomains (DRMs). Pf34 is carried into the newly invaded ring, consistent with a role for Pf34 in the formation of the parasitophorous vacuole. Pf34 is exposed to the human immune system during infection and is recognised by human immune sera collected from residents of malaria endemic areas of Vietnam and Papua New Guinea.
Figures
Similar articles
-
Conserved high activity binding peptides from the Plasmodium falciparum Pf34 rhoptry protein inhibit merozoites in vitro invasion of red blood cells.Peptides. 2010 Nov;31(11):1987-94. doi: 10.1016/j.peptides.2010.07.009. Epub 2010 Jul 21. Peptides. 2010. PMID: 20654670
-
Characterization of a membrane-associated rhoptry protein of Plasmodium falciparum.J Biol Chem. 2004 Feb 6;279(6):4648-56. doi: 10.1074/jbc.M307859200. Epub 2003 Nov 12. J Biol Chem. 2004. PMID: 14613941
-
Identification and stoichiometry of glycosylphosphatidylinositol-anchored membrane proteins of the human malaria parasite Plasmodium falciparum.Mol Cell Proteomics. 2006 Jul;5(7):1286-99. doi: 10.1074/mcp.M600035-MCP200. Epub 2006 Apr 7. Mol Cell Proteomics. 2006. PMID: 16603573
-
Erythrocyte invasion: vocabulary and grammar of the Plasmodium rhoptry.Parasitol Int. 2007 Dec;56(4):255-62. doi: 10.1016/j.parint.2007.05.003. Epub 2007 May 29. Parasitol Int. 2007. PMID: 17596999 Review.
-
Biogenesis and discharge of the rhoptries: Key organelles for entry and hijack of host cells by the Apicomplexa.Mol Microbiol. 2021 Mar;115(3):453-465. doi: 10.1111/mmi.14674. Mol Microbiol. 2021. PMID: 33368727 Review.
Cited by
-
RON12, a novel Plasmodium-specific rhoptry neck protein important for parasite proliferation.Cell Microbiol. 2014 May;16(5):657-72. doi: 10.1111/cmi.12181. Epub 2013 Aug 28. Cell Microbiol. 2014. PMID: 23937520 Free PMC article.
-
Glycosylation in malaria parasites: what do we know?Trends Parasitol. 2024 Feb;40(2):131-146. doi: 10.1016/j.pt.2023.12.006. Epub 2024 Jan 22. Trends Parasitol. 2024. PMID: 38262838 Free PMC article. Review.
-
Identification of rhoptry trafficking determinants and evidence for a novel sorting mechanism in the malaria parasite Plasmodium falciparum.PLoS Pathog. 2009 Mar;5(3):e1000328. doi: 10.1371/journal.ppat.1000328. Epub 2009 Mar 6. PLoS Pathog. 2009. PMID: 19266084 Free PMC article.
-
Identification and characterization of a novel Plasmodium falciparum merozoite apical protein involved in erythrocyte binding and invasion.PLoS One. 2008 Mar 5;3(3):e1732. doi: 10.1371/journal.pone.0001732. PLoS One. 2008. PMID: 18320051 Free PMC article.
-
A Glycosylphosphatidylinositol-Anchored Carbonic Anhydrase-Related Protein of Toxoplasma gondii Is Important for Rhoptry Biogenesis and Virulence.mSphere. 2017 May 17;2(3):e00027-17. doi: 10.1128/mSphere.00027-17. eCollection 2017 May-Jun. mSphere. 2017. PMID: 28529974 Free PMC article.
References
-
- Black CG, Wu T, Wang L, Hibbs AR, Coppel RL. Merozoite surface protein 8 of Plasmodium falciparum contains two epidermal growth factor-like domains. Mol Biochem Parasitol. 2001;114:217–226. - PubMed
-
- Black CG, Wang L, Wu T, Coppel RL. Apical location of a novel EGF-like domain-containing protein of Plasmodium falciparum. Mol Biochem Parasitol. 2003;127:59–68. - PubMed
-
- Bradley PJ, Ward C, Cheng SJ, Alexander DL, Coller S, Coombs GH, Dunn JD, Ferguson DJ, Sanderson SJ, Wastling JM, Boothroyd JC. Proteomic analysis of rhoptry organelles reveals many novel constituents for host-parasite interactions in Toxoplasma gondii. J Biol Chem. 2005;280:34245–34258. - PubMed
-
- Brown DA, London E. Structure of detergent-resistant membrane domains: does phase separation occur in biological membranes? Biochem Biophys Res Commun. 1997;240:1–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
