

Chicken heat shock protein 90 is a component of the putative cellular receptor complex of infectious bursal disease virus
- PMID: 17522206
- PMCID: PMC1951386
- DOI: 10.1128/JVI.00332-07
Chicken heat shock protein 90 is a component of the putative cellular receptor complex of infectious bursal disease virus
Abstract
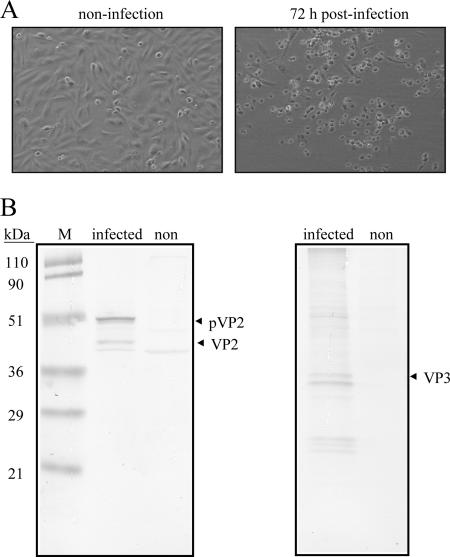
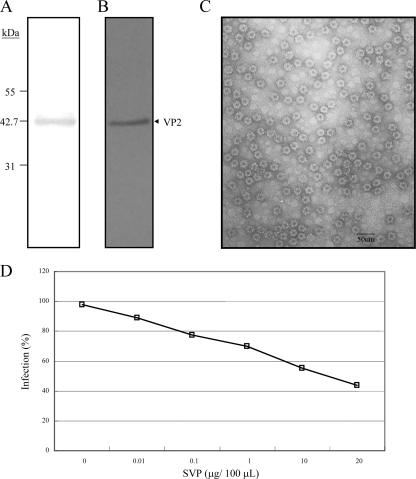
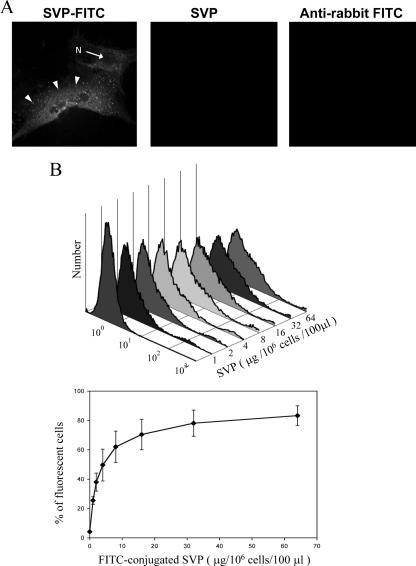
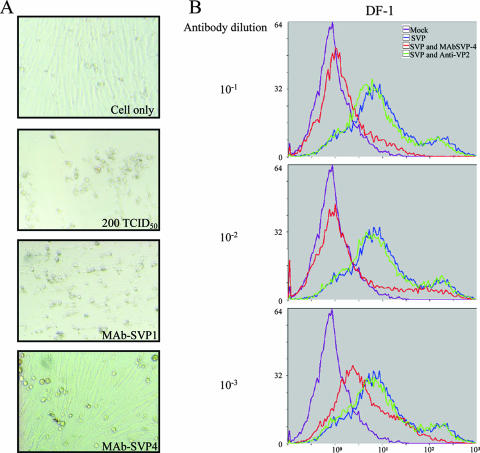
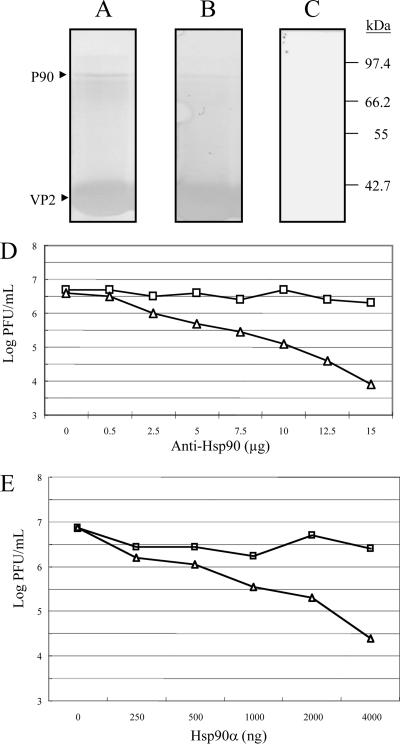
Infectious bursal disease virus (IBDV) causes a highly contagious disease in young chicks and leads to significant economic losses in the poultry industry. The capsid protein VP2 of IBDV plays an important role in virus binding and cell recognition. VP2 forms a subviral particle (SVP) with immunogenicity similar to that of the IBDV capsid. In the present study, we first showed that SVP could inhibit IBDV infection to an IBDV-susceptible cell line, DF-1 cells, in a dose-dependent manner. Second, the localizations of the SVP on the surface of DF-1 cells were confirmed by fluorescence microscopy, and the specific binding of the SVP to DF-1 cells occurred in a dose-dependent manner. Furthermore, the attachment of SVP to DF-1 cells was inhibited by an SVP-induced neutralizing monoclonal antibody against IBDV but not by denatured-VP2-induced polyclonal antibodies. Third, the cellular factors in DF-1 cells involved in the attachment of SVP were purified by affinity chromatography using SVP bound on the immobilized Ni(2+) ions. A dominant factor was identified as being chicken heat shock protein 90 (Hsp90) (cHsp90) by mass spectrometry. Results of biotinylation experiments and indirect fluorescence assays indicated that cHsp90 is located on the surface of DF-1 cells. Virus overlay protein binding assays and far-Western assays also concluded that cHsp90 interacts with IBDV and SVP, respectively. Finally, both Hsp90 and anti-Hsp90 can inhibit the infection of DF-1 cells by IBDV. Taken together, for the first time, our results suggest that cHsp90 is part of the putative cellular receptor complex essential for IBDV entry into DF-1 cells.
Figures
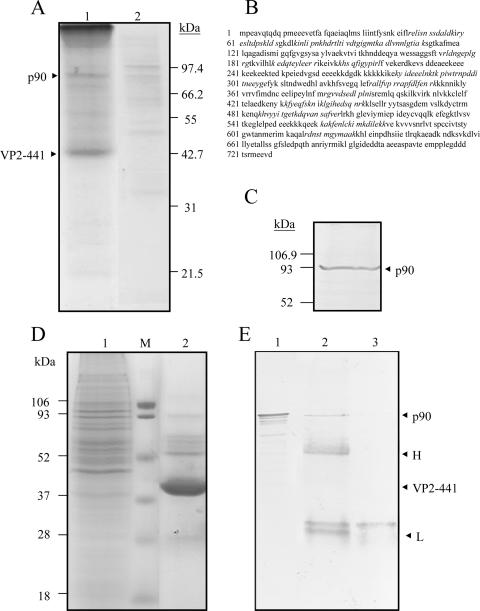
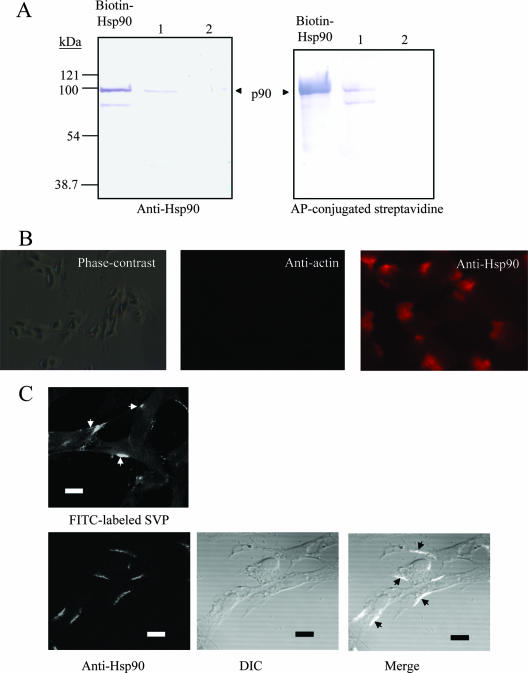
References
-
- Chen, C. S., S. Y. Suen, S. Y. Lai, G. R. Chang, T. C. Lu, M. S. Lee, and M. Y. Wang. 2005. Purification of capsid-like particles of infectious bursal disease virus (IBDV) VP2 expressed in E. coli with a metal-ion affinity membrane system. J. Virol. Methods 130:51-58. - PubMed
-
- Cheng, Y. S., M. S. Lee, S. Y. Lai, S. R. Doong, and M. Y. Wang. 2001. Separation of pure and immunoreactive virus-like particles using gel filtration chromatography following immobilized metal ion affinity chromatography. Biotechnol. Prog. 17:318-325. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
