

Replication fork velocities at adjacent replication origins are coordinately modified during DNA replication in human cells
- PMID: 17522385
- PMCID: PMC1949372
- DOI: 10.1091/mbc.e06-08-0689
Replication fork velocities at adjacent replication origins are coordinately modified during DNA replication in human cells
Abstract
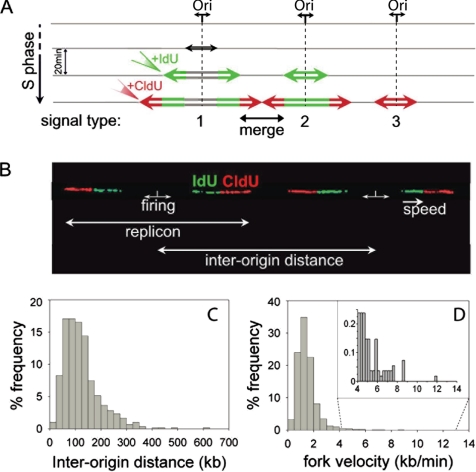
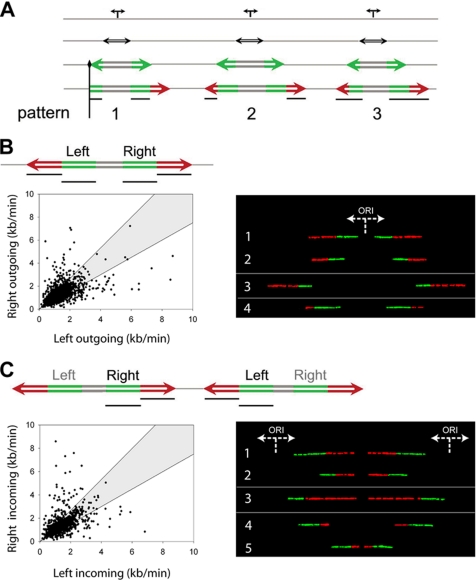
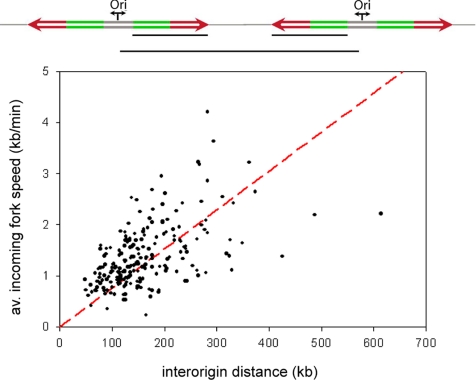
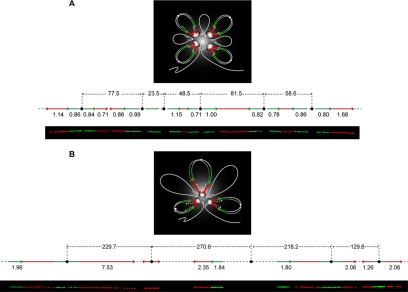
The spatial organization of replicons into clusters is believed to be of critical importance for genome duplication in higher eukaryotes, but its functional organization still remains to be fully clarified. The coordinated activation of origins is insufficient on its own to account for a timely completion of genome duplication when interorigin distances vary significantly and fork velocities are constant. Mechanisms coordinating origin distribution with fork progression are still poorly elucidated, because of technical difficulties of visualizing the process. Taking advantage of a single molecule approach, we delineated and compared the DNA replication kinetics at the genome level in human normal primary and malignant cells. Our results show that replication forks moving from one origin, as well as from neighboring origins, tend to exhibit the same velocity, although the plasticity of the replication program allows for their adaptation to variable interorigin distances. We also found that forks that emanated from closely spaced origins tended to move slower than those associated with long replicons. Taken together, our results indicate a functional role for origin clustering in the dynamic regulation of genome duplication.
Figures
References
-
- Bensimon A., Simon A., Chiffaudel A., Croquette V., Heslot F., Bensimon D. Alignment and sensitive detection of DNA by a moving interface. Science. 1994;265:2096–2098. - PubMed
-
- Berezney R., Dubey D. D., Huberman J. A. Heterogeneity of eukaryotic replicons, replicon clusters, and replication foci. Chromosoma. 2000;108:471–484. - PubMed
-
- Berezney R., Mortillaro M. J., Ma H., Wei X., Samarabandu J. The nuclear matrix: a structural milieu for genomic function. Int. Rev. Cytol. 1995;162A:1–65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
