

Use-dependent control of presynaptic calcium signalling at central synapses
- PMID: 17523936
- PMCID: PMC2375749
- DOI: 10.1111/j.1469-7580.2007.00728.x
Use-dependent control of presynaptic calcium signalling at central synapses
Abstract
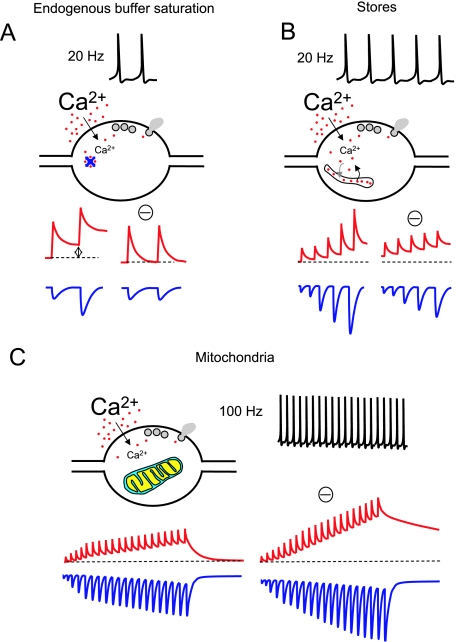
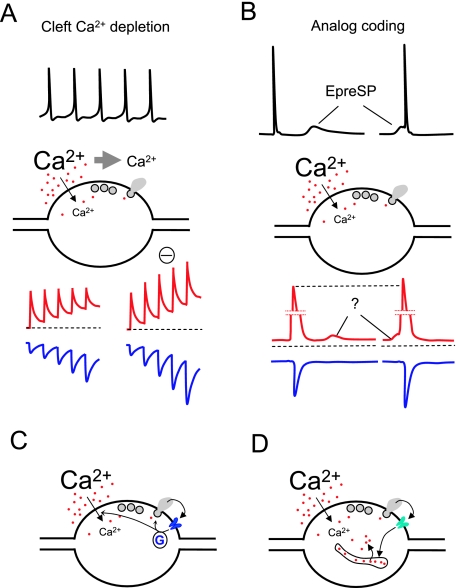
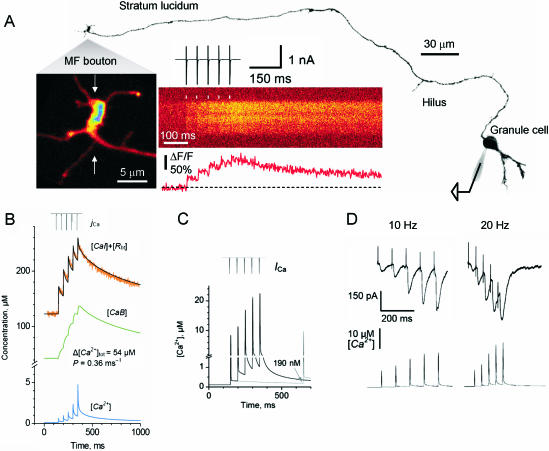
Voltage-gated Ca(2+) channels activated by action potentials evoke Ca(2+) entry into presynaptic terminals thus briefly distorting the resting Ca(2+) concentration. When this happens, a number of processes are initiated to re-establish the Ca(2+) equilibrium. During the post-spike period, the increased Ca(2+) concentration could enhance the presynaptic Ca(2+) signalling. Some of the mechanisms contributing to presynaptic Ca(2+) dynamics involve endogenous Ca(2+) buffers, Ca(2+) stores, mitochondria, the sodium-calcium exchanger, extraterminal Ca(2+) depletion and presynaptic receptors. Additionally, subthreshold presynaptic depolarization has been proposed to have an effect on release of neurotransmitters through a mechanism involving changes in resting Ca(2+). Direct evidence for the role of any of these participants in shaping the presynaptic Ca(2+) dynamics comes from direct recordings of giant presynaptic terminals and from fluorescent Ca(2+) imaging of axonal boutons. Here, some of this evidence is presented and discussed.
Figures
References
-
- Alle H, Geiger JR. Combined analog and action potential coding in hippocampal mossy fibers. Science. 2006;311:1290–1293. - PubMed
-
- Allen TJA, Noble D, Reuter H. Sodium–Calcium Exchange. Oxford: Oxford University Press; 1989.
-
- Awatramani GB, Price GD, Trussell LO. Modulation of transmitter release by presynaptic resting potential and background calcium levels. Neuron. 2005;48:109–121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
