

Optical imaging of Ca2+-evoked fluid secretion by murine nasal submucosal gland serous acinar cells
- PMID: 17525116
- PMCID: PMC2075269
- DOI: 10.1113/jphysiol.2007.131995
Optical imaging of Ca2+-evoked fluid secretion by murine nasal submucosal gland serous acinar cells
Abstract
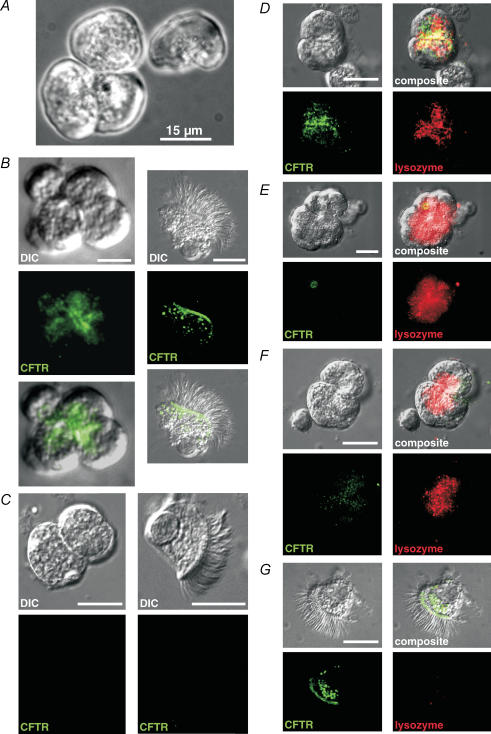
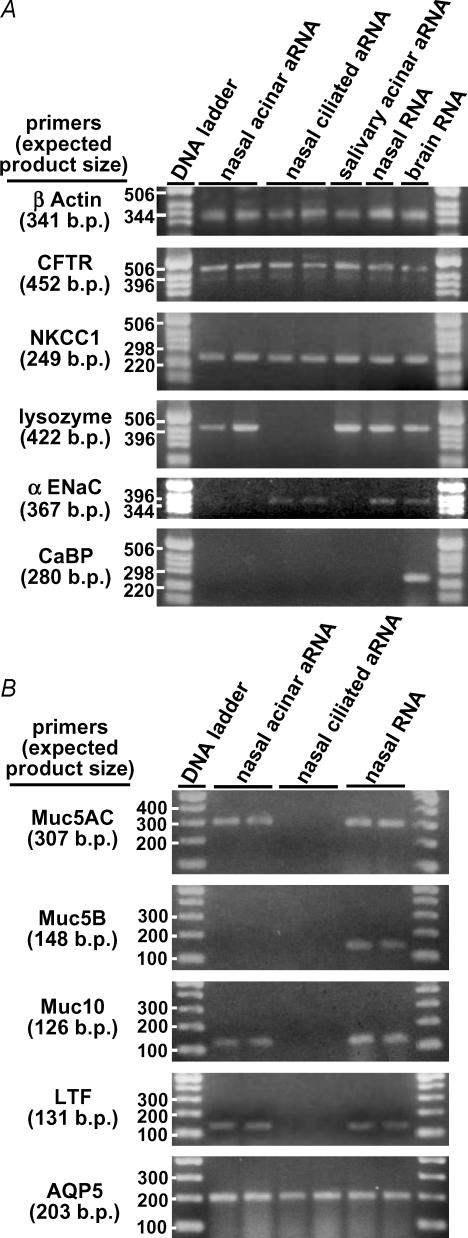
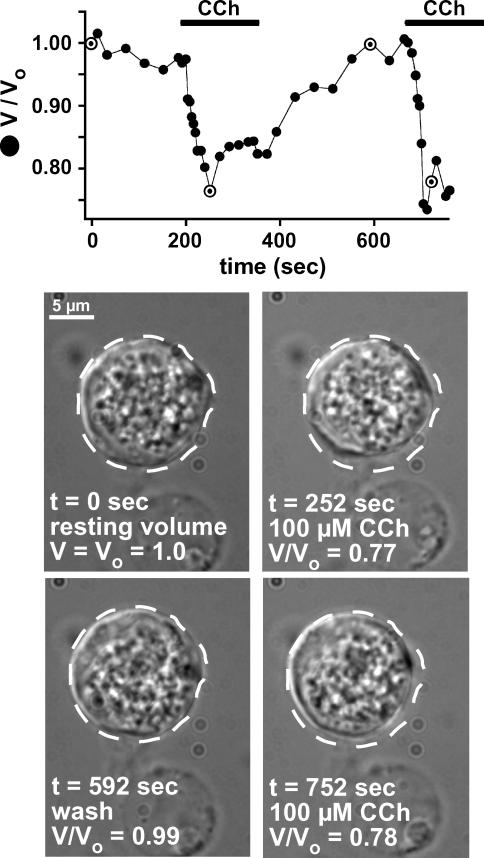
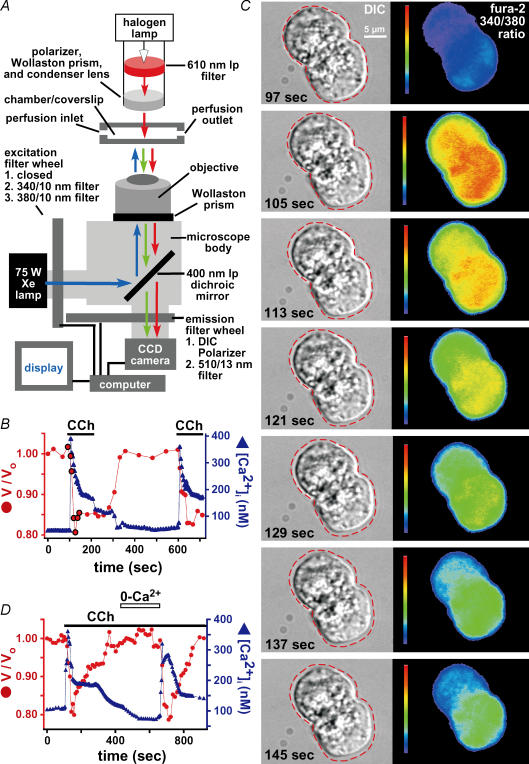
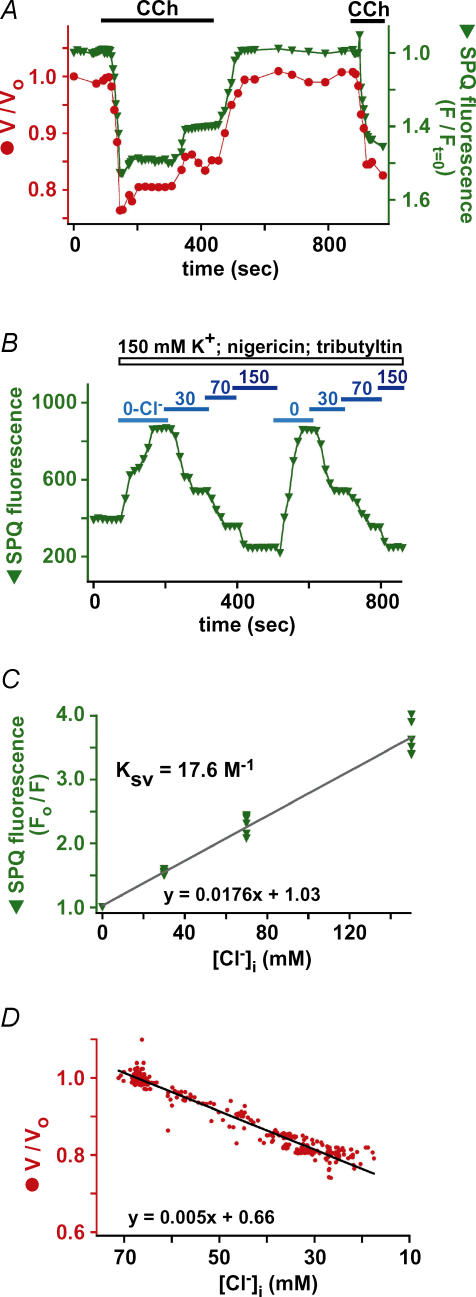
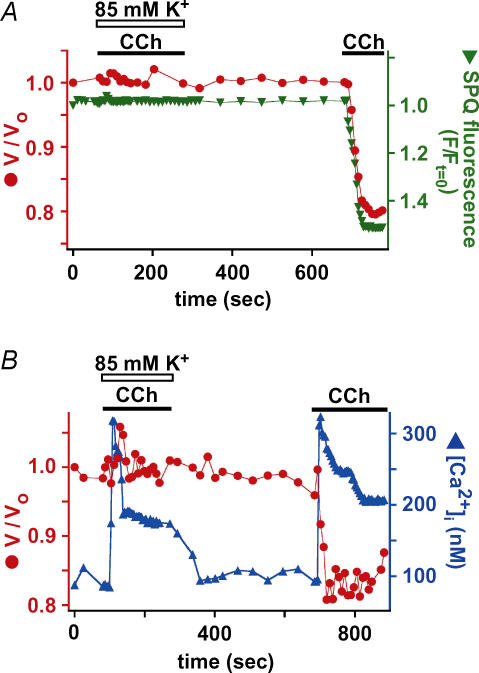
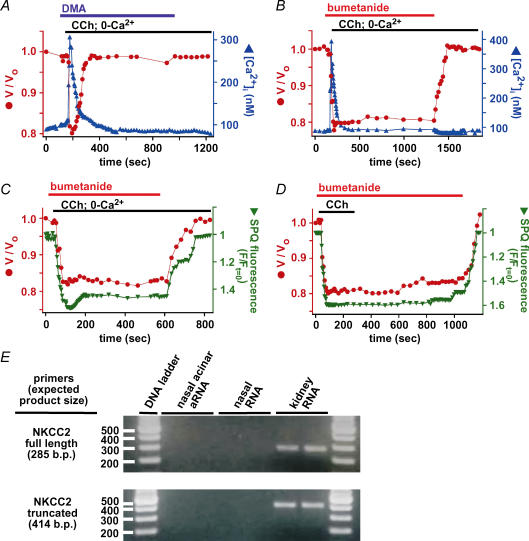
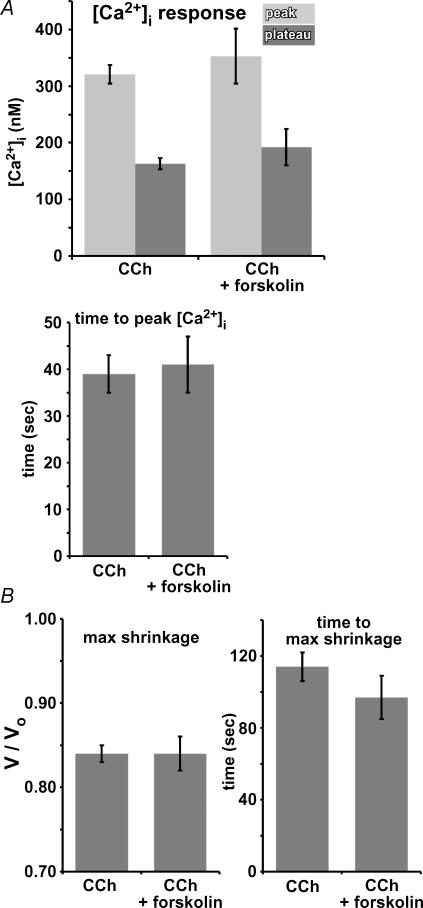
Airway submucosal glands are sites of high expression of the cystic fibrosis transmembrane conductance regulator (CFTR) Cl(-) channel and contribute to fluid homeostasis in the lung. However, the molecular mechanisms of gland ion and fluid transport are poorly defined. Here, submucosal gland serous acinar cells were isolated from murine airway, identified by immunofluorescence and gene expression profiling, and used in physiological studies. Stimulation of isolated acinar cells with carbachol (CCh), histamine or ATP was associated with marked decreases in cell volume (20 +/- 2% within 62 +/- 5 s) that were tightly correlated with increases in cytoplasmic Ca(2+) concentration ([Ca(2+)](i)) as revealed by simultaneous DIC and fluorescent indicator dye microscopy. Simultaneous imaging of cell volume and the Cl(-)-sensitive fluorophore SPQ indicated that the 20% shrinkage was associated with a fall of [Cl(-)](i) from 65 mm to 28 mm, reflecting loss of 67% of cell Cl(-) content, accompanied by parallel efflux of K(+). Upon agonist removal, [Ca(2+)](i) relaxed and the cells swelled back to resting volume via a bumetanide-sensitive Cl(-) influx pathway, likely to be NKCC1. Accordingly, agonist-induced serous acinar cell shrinkage and swelling are caused by activation of solute efflux and influx pathways, respectively, and cell volume reflects the secretory state of these cells. In contrast, elevation of cAMP failed to elicit detectible volume responses, or enhance those induced by submaximal [CCh], because the magnitude of the changes were likely to be below the threshold of detection using optical imaging. Finally, when stimulated with cholinergic or cAMP agonists, cells from mice that lacked CFTR, as well as wild-type cells treated with a CFTR inhibitor, exhibited identical rates and magnitudes of shrinkage and Cl(-) efflux compared with control cells. These results provide insights into the molecular mechanisms of salt and water secretion by lung submucosal glands, and they suggest that while murine submucosal gland fluid secretion in response to cholinergic stimulation can originate from CFTR-expressing serous acinar cells, it is not dependent upon CFTR function.
Figures




References
-
- Ballard ST, Fountain JD, Inglis SK, Corboz MR, Taylor AE. Chloride secretion across distal airway epithelium: relationship to submucosal gland distribution. Am J Physiol Lung Cell Mol Physiol. 1995;268:L526–L531. - PubMed
-
- Ballard ST, Trout L, Bebok Z, Sorscher EJ, Crews A. CFTR involvement in chloride, bicarbonate, and liquid secretion by airway submucosal glands. Am J Physiol Lung Cell Mol Physiol. 1999;277:L694–L699. - PubMed
-
- Berger JT, Voynow JA, Peters KW, Rose MC. Respiratory carcinoma cell lines. MUC genes and glycoconjugates. Am J Respir Cell Mol Biol. 1999;20:500–510. - PubMed
-
- Boucher RC. New concepts of the pathogenesis of cystic fibrosis lung disease. Eur Respir J. 2004;23:146–158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous