

Transport of truncated rhodopsin and its effects on rod function and degeneration
- PMID: 17525223
- PMCID: PMC2570206
- DOI: 10.1167/iovs.06-0035
Transport of truncated rhodopsin and its effects on rod function and degeneration
Abstract
Purpose: Most transgenic animal models of retinal degeneration caused by rhodopsin mutations express the rhodopsin transgene on a wild-type (WT) genetic background. Previous studies have demonstrated that one mechanism of retinal degeneration is rhodopsin overexpression. To study the effect of C-terminal truncation of rhodopsin without the confounding factors of overexpression, several lines of transgenic mice were generated that expressed a C-terminal rhodopsin mutation on rhodopsin-knockout backgrounds.
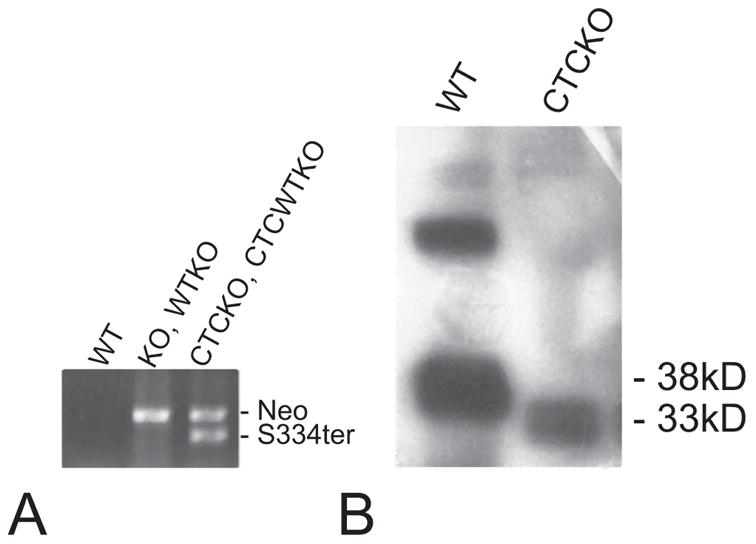
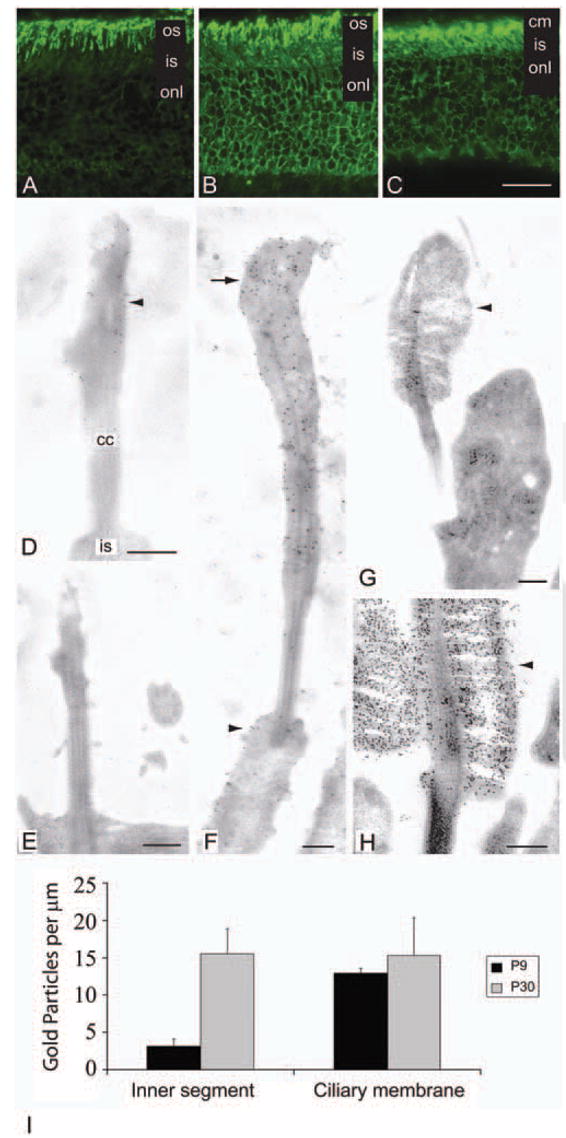
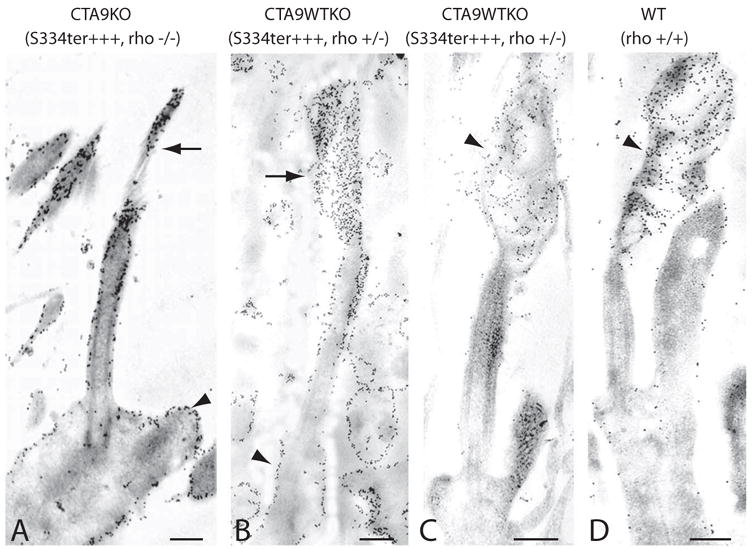
Methods: Two lines of transgenic mice, expressing different levels of C-terminal truncated rhodopsin (S334ter) were mated with heterozygous rhodopsin-knockout (rho+/-) mice to express S334ter rhodopsin on a background with reduced endogenous rhodopsin expression. S334ter mice were mated to homozygous knockout (rho-/-) mice to examine the effect of S334ter rhodopsin on a null rhodopsin background. S334ter rhodopsin expression was estimated by Western blot. Retinal function was assessed by ERG and retinal degeneration by histopathology and morphometry. C-terminal rhodopsin sorting and trafficking was examined by fluorescence immunocytochemistry with detection by electron microscope.
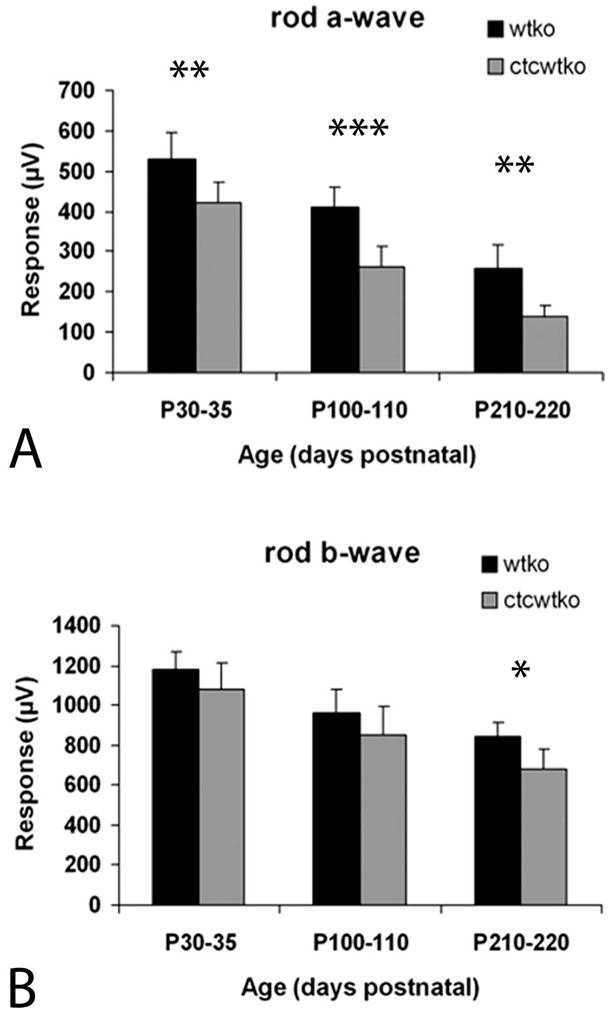
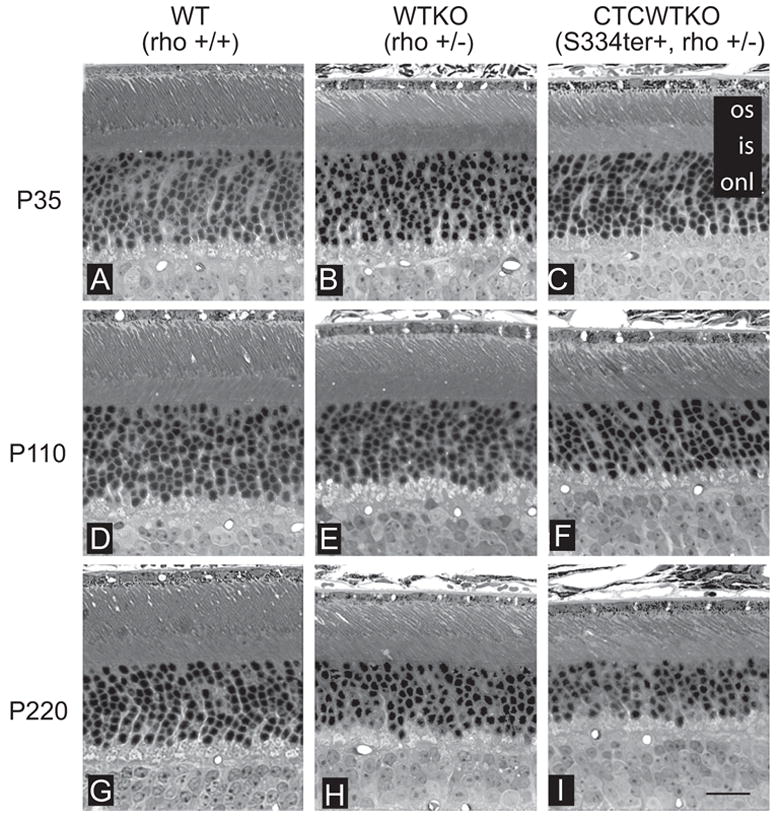
Results: Expression of S334ter truncated rhodopsin at low levels in the presence of decreased total rhodopsin in rods (S334ter, rho+/-) increased the rate of rod cell death in comparison to rho+/- littermates. In addition, S334ter rhodopsin prolonged the recovery time of the rod ERG to a light flash and diminished the a-wave amplitudes in comparison to their (rho+/-) littermates. Photoreceptors of S334ter mice on a homozygous rhodopsin-knockout background (S334ter+, rho-/-) had a fraction of mutant rhodopsin localized to the ciliary membranes.
Conclusions: Expression of S334ter rhodopsin without overexpression of total opsin in the rod photoreceptor decreased rod cell contribution to the ERG and compromised rod cell survival in adult mice. The increased cell death may be a consequence of C-terminal truncated rhodopsin mislocalization in membranes of the inner segment. Another possible pathologic mechanism is prolonged activation of phototransduction from the presence of mutant rhodopsin in the outer segment lacking the normal C-terminal binding sites for shutoff by arrestin and phosphorylation. These results suggest that rhodopsin lacking a C-terminal trafficking signal can be transported to the rod outer segment without cotransporting with full-length rhodopsin.
Figures

References
-
- Weleber RG, Gregory-Evans K. Retinitis pigmentosa and allied disorders. In: Ryan SJ, editor. Retina. St Louis: Mosby; 2001. pp. 362–470.
-
- The Retina Network (RetNet) Houston, TX: University of Texas-Houston Health Science Center; [Accessed June 10, 2005]. Available at http://www.sph.uth.tmc.edu/RetNet/disease/htm.
-
- Dryja TP, McGee TL, Hahn LB, et al. Mutations within the rhodopsin gene in patients with autosomal dominant retinitis pigmentosa. N Engl J Med. 1990;323:1302–1307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
