

The human DNA repair factor XPC-HR23B distinguishes stereoisomeric benzo[a]pyrenyl-DNA lesions
- PMID: 17525733
- PMCID: PMC1894768
- DOI: 10.1038/sj.emboj.7601730
The human DNA repair factor XPC-HR23B distinguishes stereoisomeric benzo[a]pyrenyl-DNA lesions
Abstract
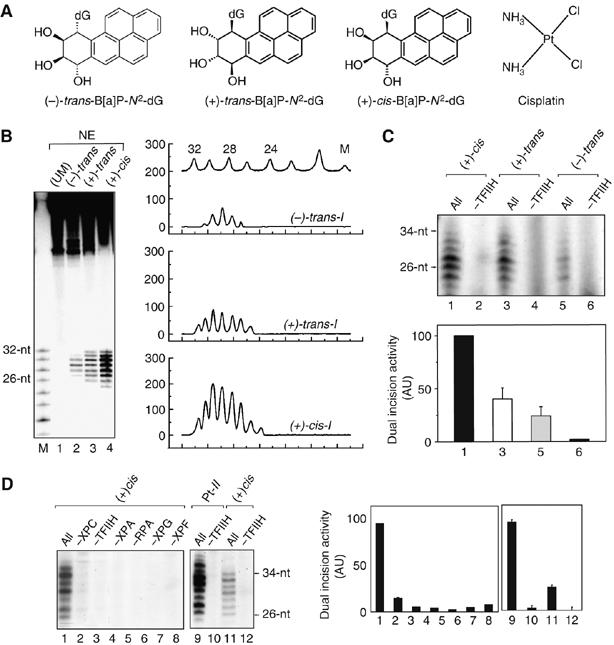
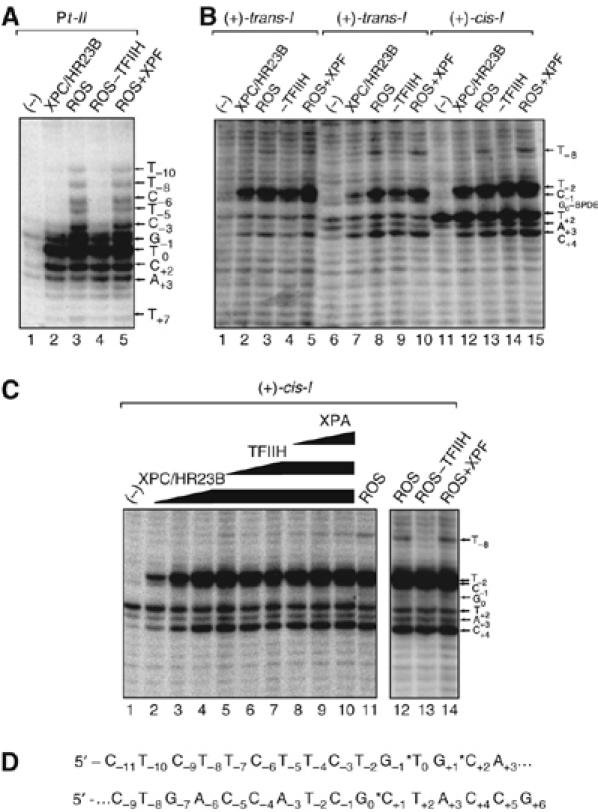
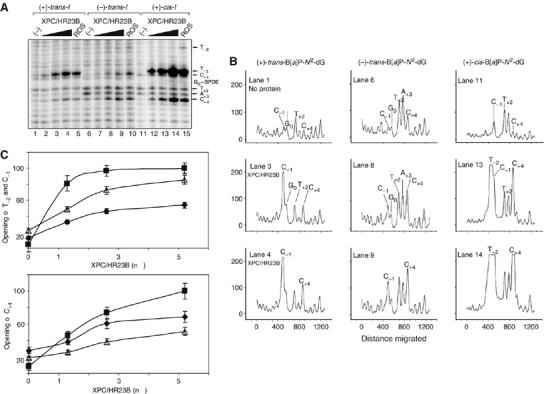
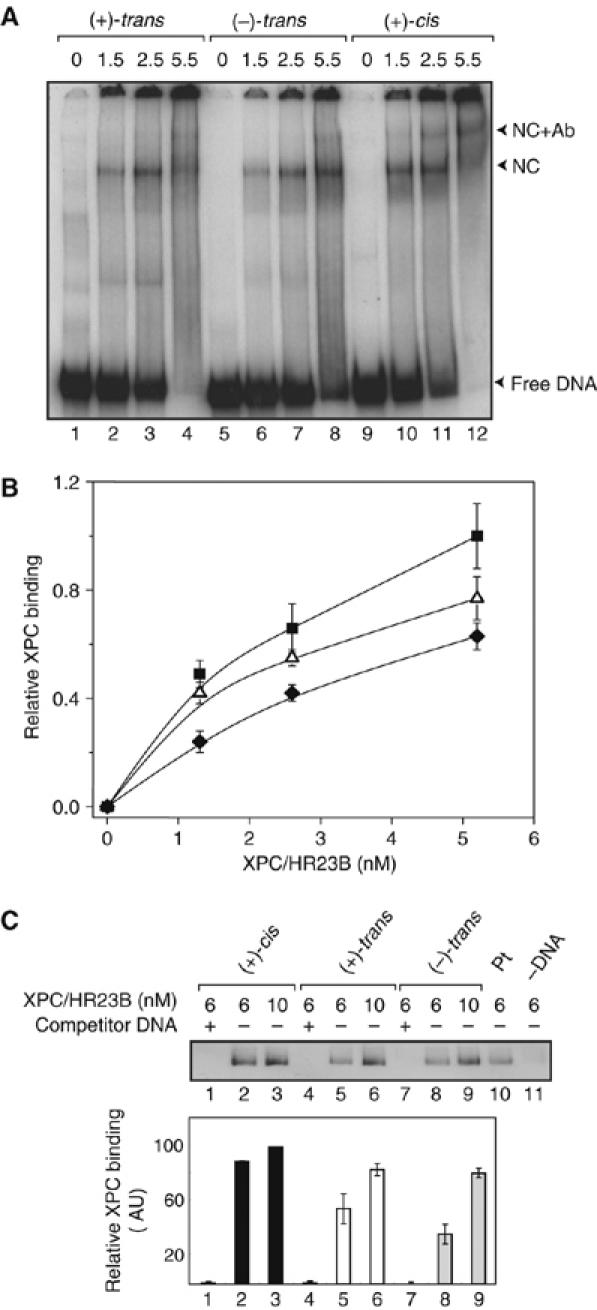
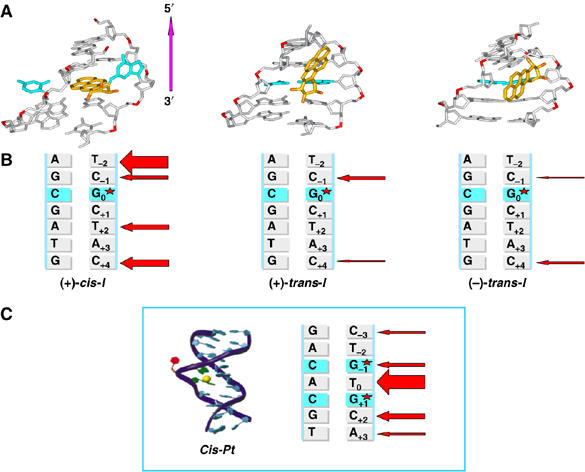
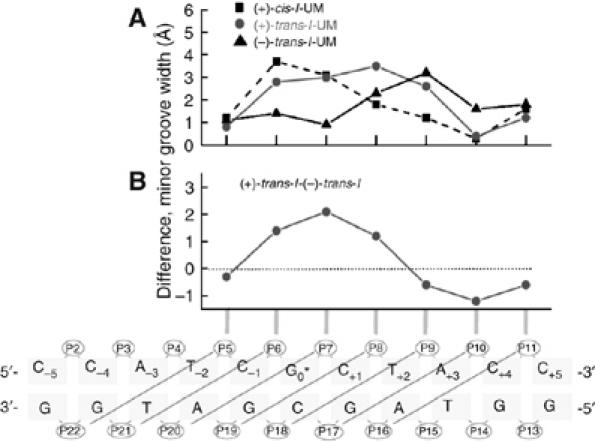
Benzo[a]pyrene (B[a]P), a known environmental pollutant and tobacco smoke carcinogen, is metabolically activated to highly tumorigenic B[a]P diol epoxide derivatives that predominantly form N(2)-guanine adducts in cellular DNA. Although nucleotide excision repair (NER) is an important cellular defense mechanism, the molecular basis of recognition of these bulky lesions is poorly understood. In order to investigate the effects of DNA adduct structure on NER, three stereoisomeric and conformationally different B[a]P-N(2)-dG lesions were site specifically incorporated into identical 135-mer duplexes and their response to purified NER factors was investigated. Using a permanganate footprinting assay, the NER lesion recognition factor XPC/HR23B exhibits, in each case, remarkably different patterns of helix opening that is also markedly distinct in the case of an intra-strand crosslinked cisplatin adduct. The different extents of helix distortions, as well as differences in the overall binding of XPC/HR23B to double-stranded DNA containing either of the three stereoisomeric B[a]P-N(2)-dG lesions, are correlated with dual incisions catalyzed by a reconstituted incision system of six purified NER factors, and by the full NER apparatus in cell-free nuclear extracts.
Figures
References
-
- Buening MK, Wislocki PG, Levin W, Yagi H, Thakker DR, Akagi H, Koreeda M, Jerina DM, Conney AH (1978) Tumorigenicity of the optical enantiomers of the diastereomeric benzo[a]pyrene 7,8-diol-9,10-epoxides in newborn mice: exceptional activity of (+)-7beta,8alpha-dihydroxy-9alpha,10alpha-epoxy-7,8,9,10-tetrahydrobenzo[a ]pyrene. Proc Natl Acad Sci USA 75: 5358–5361 - PMC - PubMed
-
- Buterin T, Meyer C, Giese B, Naegeli H (2005) DNA quality control by conformational readout on the undamaged strand of the double helix. Chem Biol 12: 913–922 - PubMed
-
- Cheatham TE, Cieplak P, Kollman PA (1999) A modified version of the Cornell et al force field with improved sugar pucker phases and helical repeat. J Biomol Struct Dyn 16: 845–862 - PubMed
-
- Conney AH (1982) Induction of microsomal enzymes by foreign chemicals and carcinogenesis by polycyclic aromatic hydrocarbons. Cancer Res 42: 4875–4917 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
