

C-terminal regions of topoisomerase IIalpha and IIbeta determine isoform-specific functioning of the enzymes in vivo
- PMID: 17526531
- PMCID: PMC1920234
- DOI: 10.1093/nar/gkm102
C-terminal regions of topoisomerase IIalpha and IIbeta determine isoform-specific functioning of the enzymes in vivo
Abstract
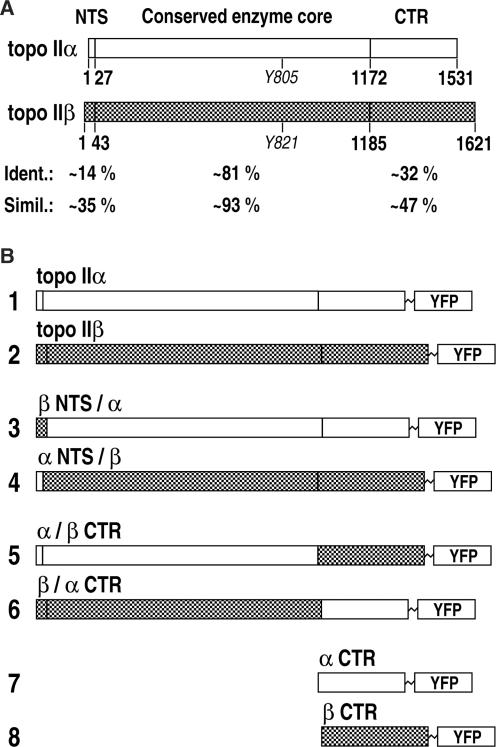
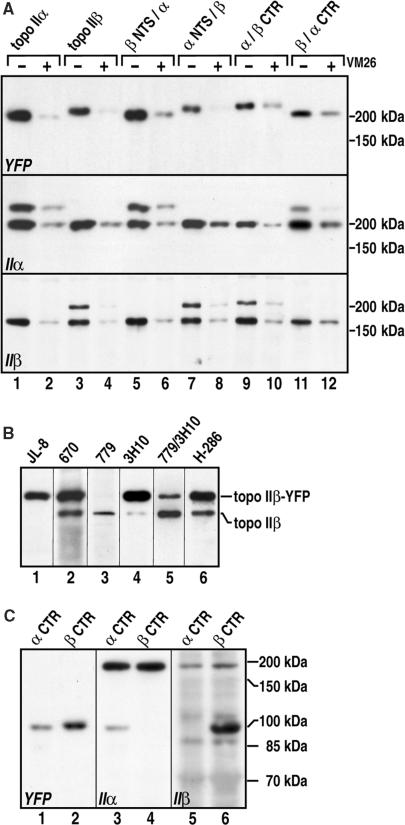
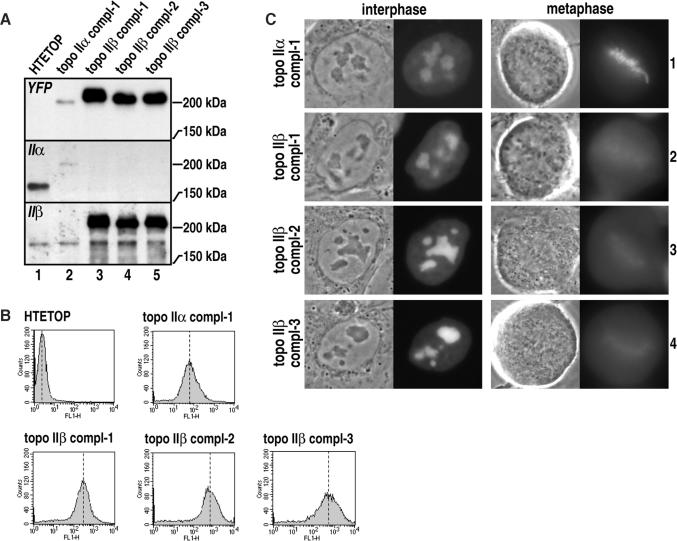
Topoisomerase II removes supercoils and catenanes generated during DNA metabolic processes such as transcription and replication. Vertebrate cells express two genetically distinct isoforms (alpha and beta) with similar structures and biochemical activities but different biological roles. Topoisomerase IIalpha is essential for cell proliferation, whereas topoisomerase IIbeta is required only for aspects of nerve growth and brain development. To identify the structural features responsible for these differences, we exchanged the divergent C-terminal regions (CTRs) of the two human isoforms (alpha 1173-1531 and beta 1186-1621) and tested the resulting hybrids for complementation of a conditional topoisomerase IIalpha knockout in human cells. Proliferation was fully supported by all enzymes bearing the alpha CTR. The alpha CTR also promoted chromosome binding of both enzyme cores, and was by itself chromosome-bound, suggesting a role in enzyme targeting during mitosis. In contrast, enzymes bearing the beta CTR supported proliferation only rarely and when expressed at unusually high levels. A similar analysis of the divergent N-terminal regions (alpha 1-27 and beta 1-43) revealed no role in isoform-specific functions. Our results show that it is the CTRs of human topoisomerase II that determine their isoform-specific functions in proliferating cells. They also indicate persistence of some functional redundancy between the two isoforms.
Figures


References
-
- Uemura T, Ohkura H, Adachi Y, Morino K, Shiozaki K, Yanagida M. DNA topoisomerase II is required for condensation and separation of mitotic chromosomes in S. pombe. Cell. 1987;50:917–925. - PubMed
-
- Holm C, Goto T, Wang JC, Botstein D. DNA topoisomerase II is required at the time of mitosis in yeast. Cell. 1985;41:553–563. - PubMed
-
- Sundin O, Varshavsky A. Arrest of segregation leads to accumulation of highly intertwined catenated dimers: dissection of the final stages of SV40 DNA replication. Cell. 1981;25:659–669. - PubMed
-
- Liu LF. DNA topoisomerase poisons as antitumor drugs. Annu. Rev. Biochem. 1989;58:351–375. - PubMed
-
- Kingma PS, Osheroff N. The response of eukaryotic topoisomerases to DNA damage. Biochim. Biophys. Acta. 1998;1400:223–232. - PubMed
