

The two-component regulatory system senX3-regX3 regulates phosphate-dependent gene expression in Mycobacterium smegmatis
- PMID: 17526710
- PMCID: PMC1951828
- DOI: 10.1128/JB.00190-07
The two-component regulatory system senX3-regX3 regulates phosphate-dependent gene expression in Mycobacterium smegmatis
Abstract
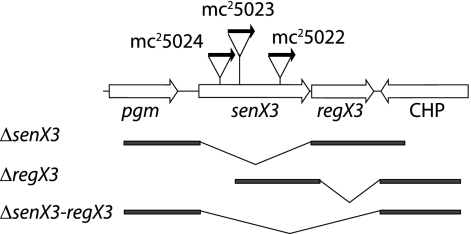
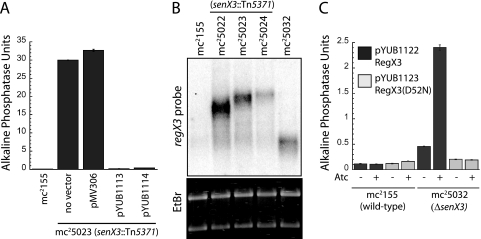
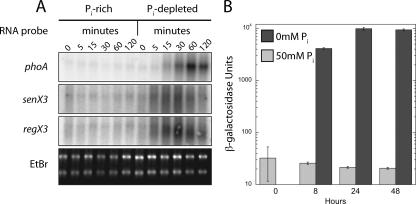
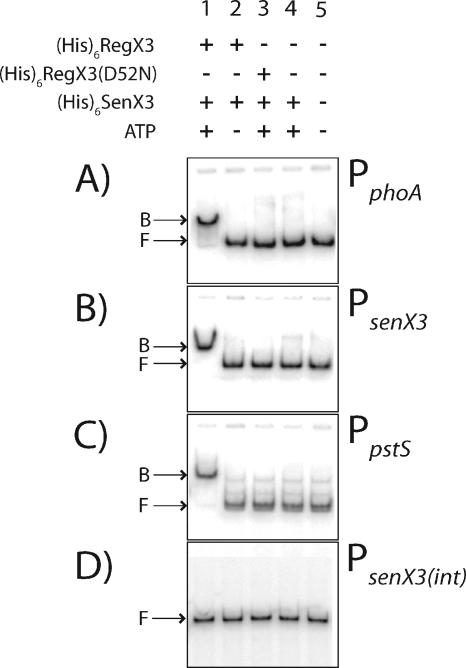
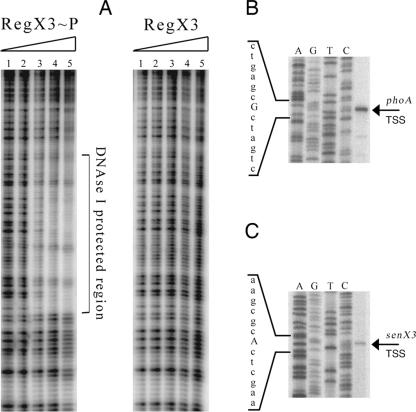
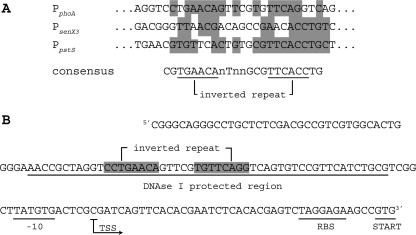
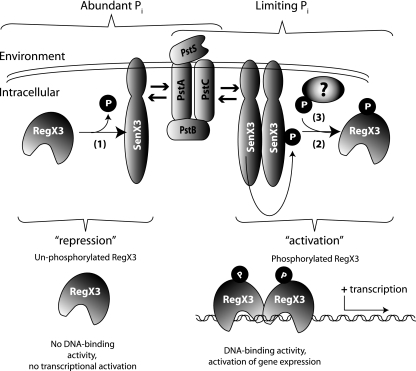
Phosphate import is required for the growth of mycobacteria and is regulated by environmental inorganic phosphate (P(i)) concentrations, although the mechanism of this regulation has not been characterized. The expression of genes involved in P(i) acquisition is frequently regulated by two-component regulatory systems (2CRs) consisting of a sensor histidine kinase and a DNA-binding response regulator. In this work, we have identified the senX3-regX3 2CR as a P(i)-dependent regulator of genes involved in phosphate acquisition in Mycobacterium smegmatis. Characterization of senX3 mutants with different PhoA phenotypes suggests a dual role for SenX3 as a phosphatase or a phosphodonor for the response regulator RegX3, depending upon P(i) availability. Expression of PhoA activity required phosphorylation of RegX3, consistent with a role for phosphorylated RegX3 (RegX3 approximately P) as a transcriptional activator of phoA. Furthermore, purified RegX3 approximately P bound to promoter sequences from phoA, senX3, and the high-affinity phosphate transporter component pstS, demonstrating direct transcriptional control of all three genes. DNase I footprinting and primer extension analyses have further defined the DNA-binding region and transcriptional start site within the phoA promoter. A DNA motif consisting of an inverted repeat was identified in each of the promoters bound by RegX3 approximately P. Based upon our findings, we propose a model for P(i)-regulated gene expression mediated by SenX3-RegX3 in mycobacteria.
Figures
References
-
- Andersen, A. B., L. Ljungqvist, and M. Olsen. 1990. Evidence that protein antigen b of Mycobacterium tuberculosis is involved in phosphate metabolism. J. Gen. Microbiol. 136:477-480. - PubMed
-
- Barletta, R. G., D. D. Kim, S. B. Snapper, B. R. Bloom, and W. R. Jacobs, Jr. 1992. Identification of expression signals of the mycobacteriophages Bxb1, L1 and TM4 using the Escherichia-Mycobacterium shuttle plasmids pYUB75 and pYUB76 designed to create translational fusions to the lacZ gene. J. Gen. Microbiol. 138:23-30. - PubMed
-
- Braibant, M., and J. Content. 2001. The cell surface associated phosphatase activity of Mycobacterium bovis BCG is not regulated by environmental inorganic phosphate. FEMS Microbiol. Lett. 195:121-126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
