

Specificity and complexity of the Caenorhabditis elegans innate immune response
- PMID: 17526726
- PMCID: PMC1952075
- DOI: 10.1128/MCB.02070-06
Specificity and complexity of the Caenorhabditis elegans innate immune response
Abstract
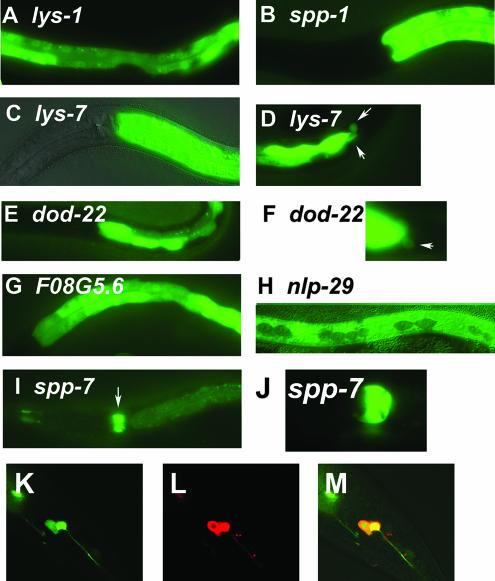
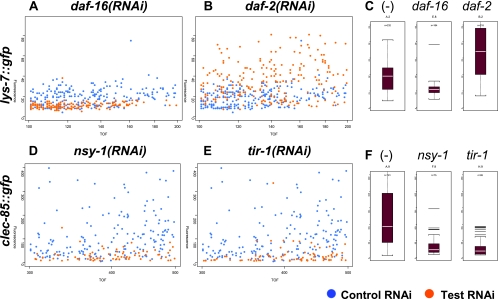
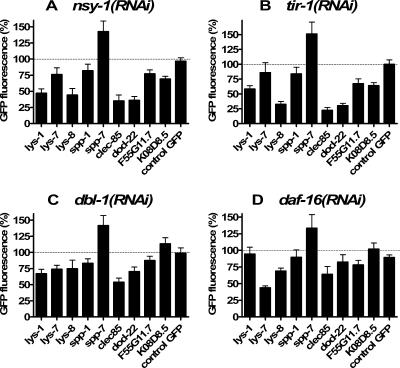
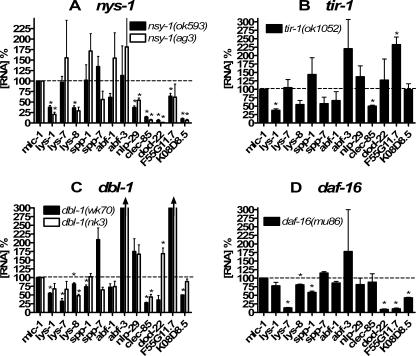
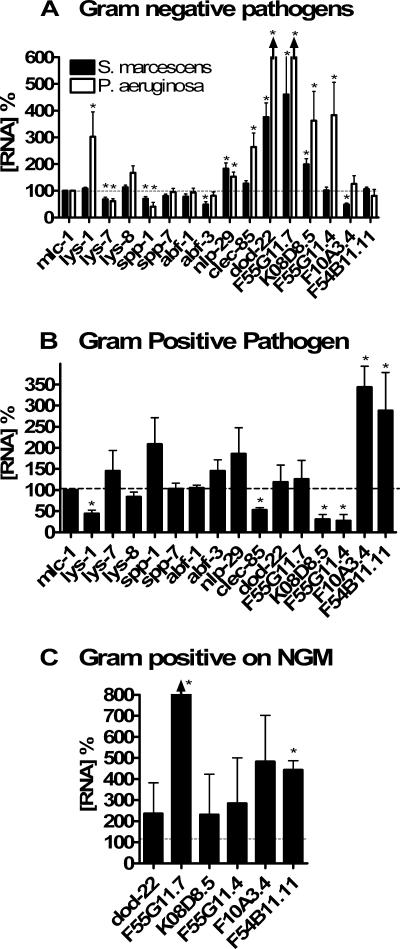
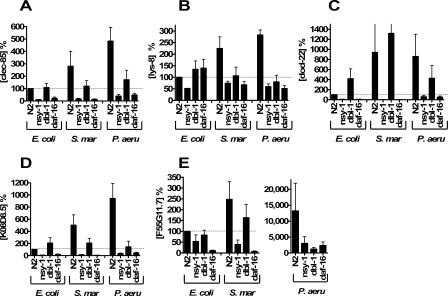
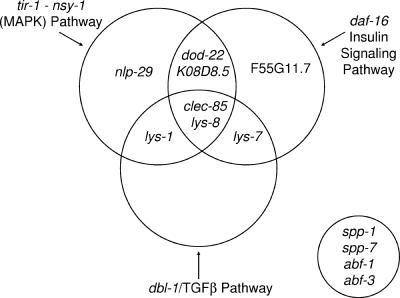
In response to infection, Caenorhabditis elegans produces an array of antimicrobial proteins. To understand the C. elegans immune response, we have investigated the regulation of a large, representative sample of candidate antimicrobial genes. We found that all these putative antimicrobial genes are expressed in tissues exposed to the environment, a position from which they can ward off infection. Using RNA interference to inhibit the function of immune signaling pathways in C. elegans, we found that different immune response pathways regulate expression of distinct but overlapping sets of antimicrobial genes. We also show that different bacterial pathogens regulate distinct but overlapping sets of antimicrobial genes. The patterns of genes induced by pathogens do not coincide with any single immune signaling pathway. Thus, even in this simple model system for innate immunity, striking specificity and complexity exist in the immune response. The unique patterns of antimicrobial gene expression observed when C. elegans is exposed to different pathogens or when different immune signaling pathways are perturbed suggest that a large set of yet to be identified pathogen recognition receptors (PRRs) exist in the nematode. These PRRs must interact in a complicated fashion to induce a unique set of antimicrobial genes. We also propose the existence of an "antimicrobial fingerprint," which will aid in assigning newly identified C. elegans innate immunity genes to known immune signaling pathways.
Figures
References
-
- Aballay, A., E. Drenkard, L. R. Hilbun, and F. M. Ausubel. 2003. Caenorhabditis elegans innate immune response triggered by Salmonella enterica requires intact LPS and is mediated by a MAPK signaling pathway. Curr. Biol. 13:47-52. - PubMed
-
- Ahringer, J. (ed.). 6 April 2006, posting date. Reverse genetics. In The C. elegans Research Community (ed.), WormBook. http://www.wormbook.org.
-
- Alegado, R. A., M. C. Campbell, W. C. Chen, S. S. Slutz, and M. W. Tan. 2003. Characterization of mediators of microbial virulence and innate immunity using the Caenorhabditis elegans host-pathogen model. Cell. Microbiol. 5:435-444. - PubMed
-
- Banyai, L., and L. Patthy. 1998. Amoebapore homologs of Caenorhabditis elegans. Biochim. Biophys. Acta 1429:259-264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases