

Cytoplasmic remodeling of erythrocyte raft lipids during infection by the human malaria parasite Plasmodium falciparum
- PMID: 17526861
- PMCID: PMC1976375
- DOI: 10.1182/blood-2007-04-083873
Cytoplasmic remodeling of erythrocyte raft lipids during infection by the human malaria parasite Plasmodium falciparum
Abstract
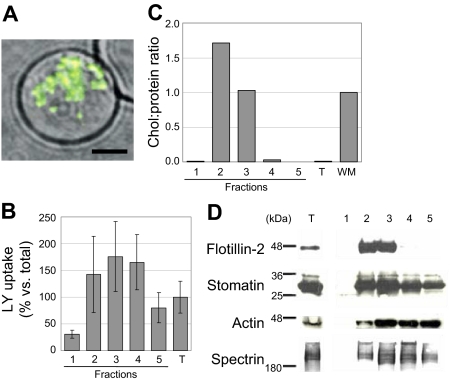
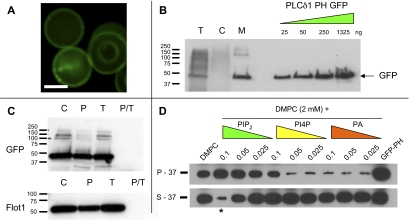
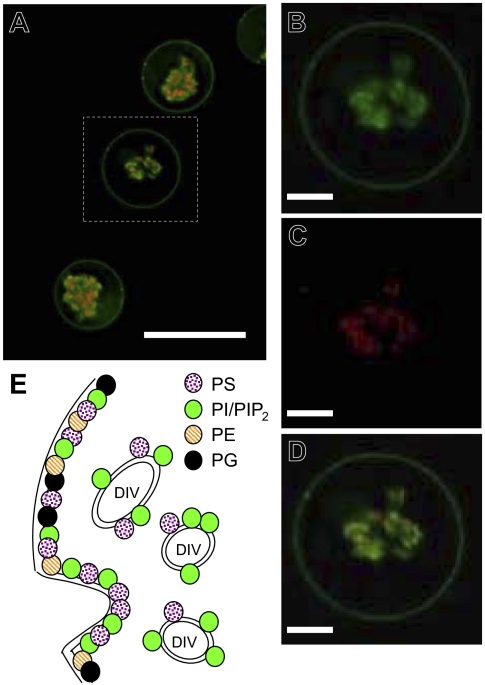
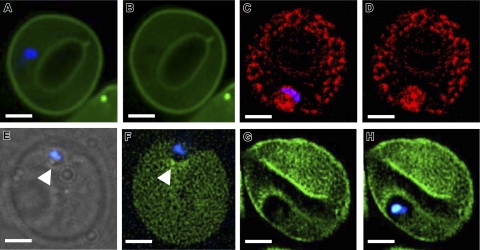
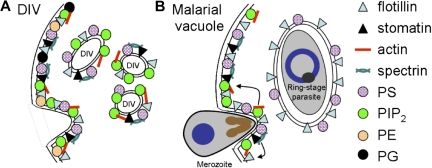
Studies of detergent-resistant membrane (DRM) rafts in mature erythrocytes have facilitated identification of proteins that regulate formation of endovacuolar structures such as the parasitophorous vacuolar membrane (PVM) induced by the malaria parasite Plasmodium falciparum. However, analyses of raft lipids have remained elusive because detergents interfere with lipid detection. Here, we use primaquine to perturb the erythrocyte membrane and induce detergent-free buoyant vesicles, which are enriched in cholesterol and major raft proteins flotillin and stomatin and contain low levels of cytoskeleton, all characteristics of raft microdomains. Lipid mass spectrometry revealed that phosphatidylethanolamine and phosphatidylglycerol are depleted in endovesicles while phosphoinositides are highly enriched, suggesting raft-based endovesiculation can be achieved by simple (non-receptor-mediated) mechanical perturbation of the erythrocyte plasma membrane and results in sorting of inner leaflet phospholipids. Live-cell imaging of lipid-specific protein probes showed that phosphatidylinositol (4,5) bisphosphate (PIP(2)) is highly concentrated in primaquine-induced vesicles, confirming that it is an erythrocyte raft lipid. However, the malarial PVM lacks PIP(2), although another raft lipid, phosphatidylserine, is readily detected. Thus, different remodeling/sorting of cytoplasmic raft phospholipids may occur in distinct endovacuoles. Importantly, erythrocyte raft lipids recruited to the invasion junction by mechanical stimulation may be remodeled by the malaria parasite to establish blood-stage infection.
Figures
Similar articles
-
Erythrocyte detergent-resistant membrane proteins: their characterization and selective uptake during malarial infection.Blood. 2004 Mar 1;103(5):1920-8. doi: 10.1182/blood-2003-09-3165. Epub 2003 Oct 30. Blood. 2004. PMID: 14592818
-
Identification of a stomatin orthologue in vacuoles induced in human erythrocytes by malaria parasites. A role for microbial raft proteins in apicomplexan vacuole biogenesis.J Biol Chem. 2003 Nov 28;278(48):48413-21. doi: 10.1074/jbc.M307266200. Epub 2003 Sep 10. J Biol Chem. 2003. PMID: 12968029
-
Lipid rafts and malaria parasite infection of erythrocytes.Mol Membr Biol. 2006 Jan-Feb;23(1):81-8. doi: 10.1080/09687860500473440. Mol Membr Biol. 2006. PMID: 16611583 Review.
-
Trafficking of malarial proteins to the host cell cytoplasm and erythrocyte surface membrane involves multiple pathways.J Cell Biol. 1992 Dec;119(6):1481-95. doi: 10.1083/jcb.119.6.1481. J Cell Biol. 1992. PMID: 1469045 Free PMC article.
-
Protein and lipid trafficking induced in erythrocytes infected by malaria parasites.Cell Microbiol. 2002 Jul;4(7):383-95. doi: 10.1046/j.1462-5822.2002.00204.x. Cell Microbiol. 2002. PMID: 12102685 Review.
Cited by
-
Erythropoietin receptor signaling is membrane raft dependent.PLoS One. 2012;7(4):e34477. doi: 10.1371/journal.pone.0034477. Epub 2012 Apr 3. PLoS One. 2012. PMID: 22509308 Free PMC article.
-
Hemozoin: a waste product after heme detoxification?Parasit Vectors. 2025 Mar 4;18(1):83. doi: 10.1186/s13071-025-06699-x. Parasit Vectors. 2025. PMID: 40038801 Free PMC article.
-
Plasmodium falciparum infection-induced changes in erythrocyte membrane proteins.Parasitol Res. 2012 Feb;110(2):545-56. doi: 10.1007/s00436-011-2521-2. Epub 2011 Jul 9. Parasitol Res. 2012. PMID: 21744020
-
Physicochemical Aspects of the Plasmodium chabaudi-Infected Erythrocyte.Biomed Res Int. 2015;2015:642729. doi: 10.1155/2015/642729. Epub 2015 Oct 18. Biomed Res Int. 2015. PMID: 26557685 Free PMC article.
-
4D analysis of malaria parasite invasion offers insights into erythrocyte membrane remodeling and parasitophorous vacuole formation.Nat Commun. 2021 Jun 15;12(1):3620. doi: 10.1038/s41467-021-23626-7. Nat Commun. 2021. PMID: 34131147 Free PMC article.
References
-
- Dvorak JA, Miller LH, Whitehouse WC, Shiroishi T. Invasion of erythrocytes by malaria merozoites. Science. 1975;187:748–750. - PubMed
-
- Holder AA, Blackman MJ, Borre M, et al. Malaria parasites and erythrocyte invasion. Biochem Soc Trans. 1994;22:291–295. - PubMed
-
- Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria. Nature. 2002;415:673–679. - PubMed
-
- Murphy SC, Samuel BU, Harrison T, et al. Erythrocyte detergent-resistant membrane proteins: their characterization and selective uptake during malarial infection. Blood. 2004;103:1920–1928. - PubMed
-
- Samuel BU, Mohandas N, Harrison T, et al. The role of cholesterol and glycosylphosphatidylinositol-anchored proteins of erythrocyte rafts in regulating raft protein content and malarial infection. J Biol Chem. 2001;276:29319–29329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
