

Differential stability of dimeric and monomeric cytochrome c oxidase exposed to elevated hydrostatic pressure
- PMID: 17530783
- PMCID: PMC2561924
- DOI: 10.1021/bi700548a
Differential stability of dimeric and monomeric cytochrome c oxidase exposed to elevated hydrostatic pressure
Abstract
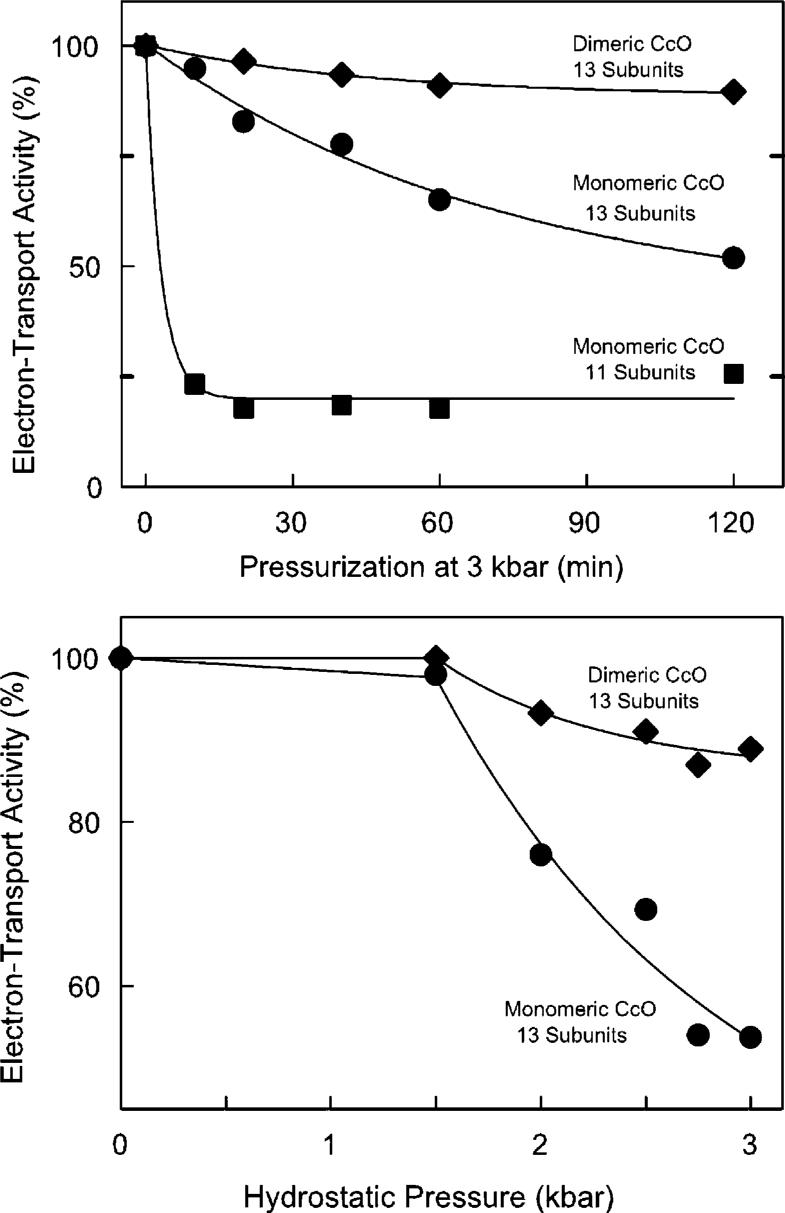
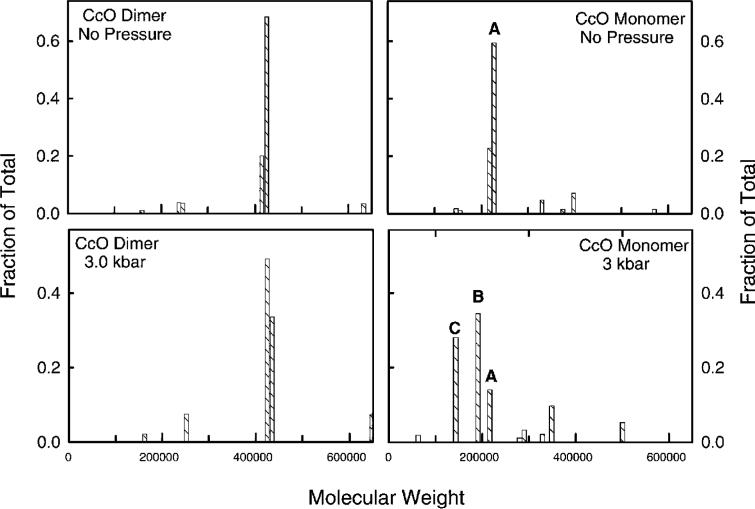
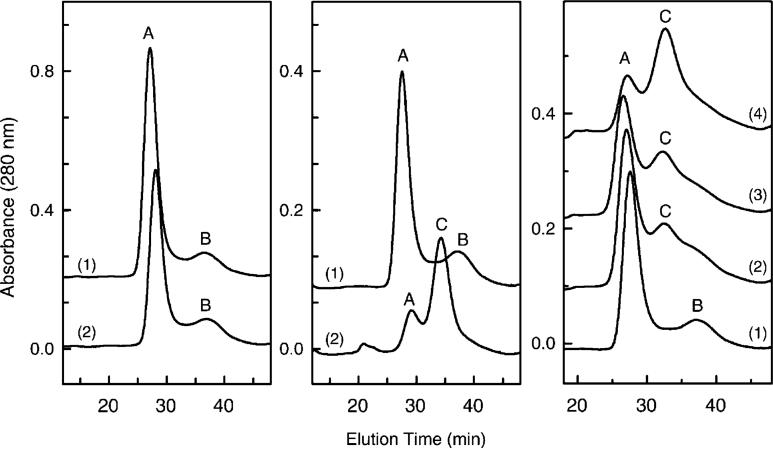
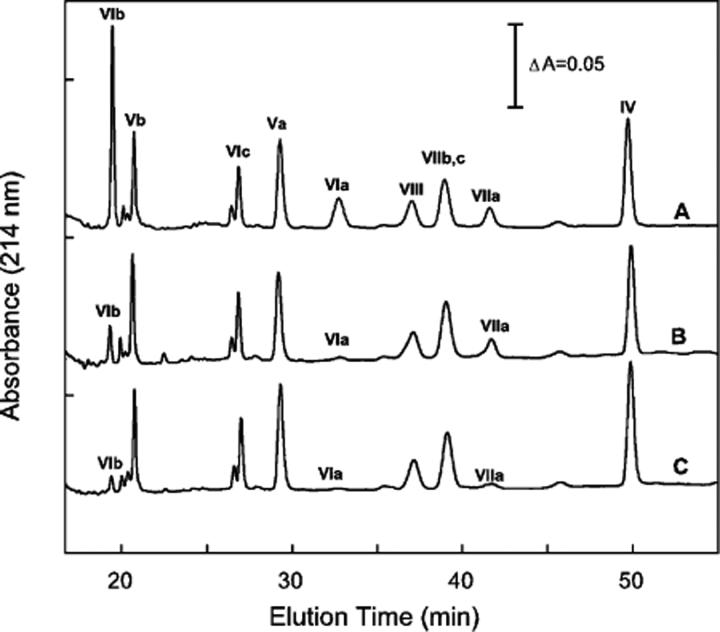
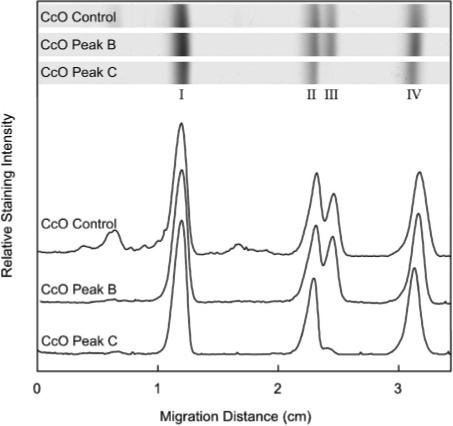
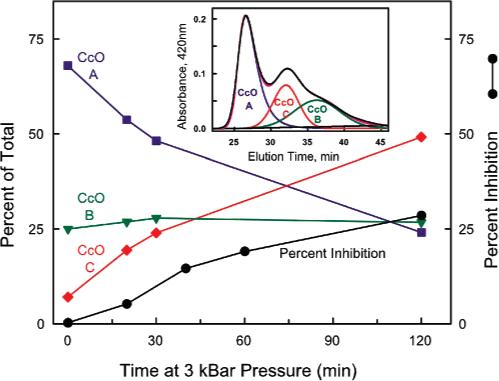
Detergent-solubilized dimeric and monomeric cytochrome c oxidase (CcO) have significantly different quaternary stability when exposed to 2-3 kbar of hydrostatic pressure. Dimeric, dodecyl maltoside-solubilized cytochrome c oxidase is very resistant to elevated hydrostatic pressure with almost no perturbation of its quaternary structure or functional activity after release of pressure. In contrast to the stability of dimeric CcO, 3 kbar of hydrostatic pressure triggers multiple structural and functional alterations within monomeric cytochrome c oxidase. The perturbations are either irreversible or slowly reversible since they persist after the release of high pressure. Therefore, standard biochemical analytical procedures could be used to quantify the pressure-induced changes after the release of hydrostatic pressure. The electron transport activity of monomeric cytochrome c oxidase decreases by as much as 60% after exposure to 3 kbar of hydrostatic pressure. The irreversible loss of activity occurs in a time- and pressure-dependent manner. Coincident with the activity loss is a sequential dissociation of four subunits as detected by sedimentation velocity, high-performance ion-exchange chromatography, and reversed-phase and SDS-PAGE subunit analysis. Subunits VIa and VIb are the first to dissociate followed by subunits III and VIIa. Removal of subunits VIa and VIb prior to pressurization makes the resulting 11-subunit form of CcO even more sensitive to elevated hydrostatic pressure than monomeric CcO containing all 13 subunits. However, dimeric CcO, in which the association of VIa and VIb is stabilized, is not susceptible to pressure-induced inactivation. We conclude that dissociation of subunit III and/or VIIa must be responsible for pressure-induced inactivation of CcO since VIa and VIb can be removed from monomeric CcO without significant activity loss. These results are the first to clearly demonstrate an important structural role for the dimeric form of cytochrome c oxidase, i.e., stabilization of its quaternary structure.
Figures
References
-
- Tsukihara T, Aoyama H, Yamashita E, Tomizaki T, Yamaguchi H, Shinzawa-Itoh K, Nakashima R, Yaono R, Yoshikawa S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 Å. Science. 1996;272:1136–1144. - PubMed
-
- Kadenbach B, Jarausch J, Hartmann R, Merle P. Separation of mammalian cytochrome c oxidase into 13 polypeptides by sodium dodecyl sulfate-gel electrophoresis procedure. Anal. Biochem. 1983;129:517–521. - PubMed
-
- Robinson NC. Functional binding of cardiolipin to cytochrome c oxidase. J. Bioenerg. Biomembr. 1993;25:153–163. - PubMed
-
- Suarez MD, Revzin A, Narlock R, Kempner ES, Thompson DA, Ferguson-Miller S. The functional and physical form of mammalian cytochrome c oxidase determined by gel filtration, radiation inactivation, and sedimentation equilibrium analysis. J. Biol. Chem. 1984;259:13791–13799. - PubMed
-
- Robinson NC, Talbert L. Triton X-100 induced dissociation of beef heart cytochrome c oxidase into monomers. Biochemistry. 1986;25:2328–2335. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
