

The emergence of alternative 3' and 5' splice site exons from constitutive exons
- PMID: 17530917
- PMCID: PMC1876488
- DOI: 10.1371/journal.pcbi.0030095
The emergence of alternative 3' and 5' splice site exons from constitutive exons
Abstract
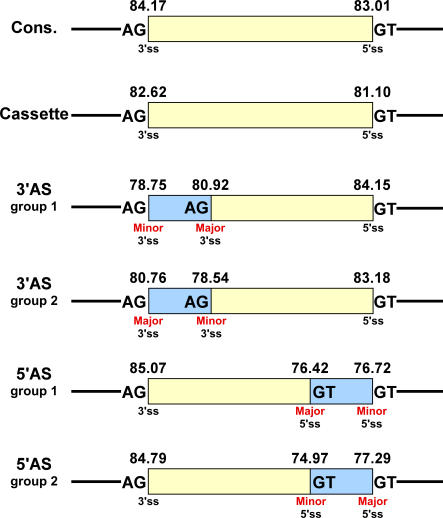
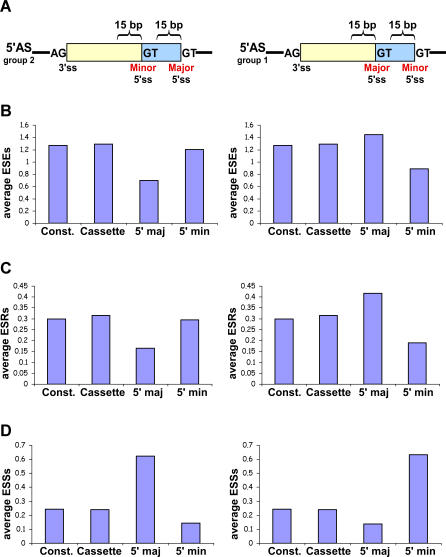
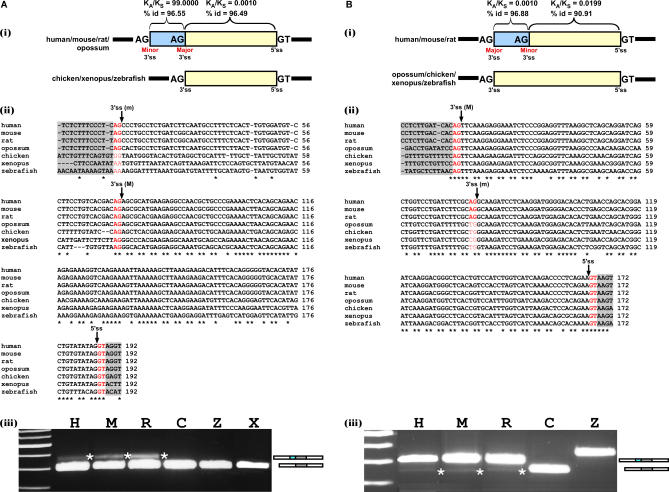
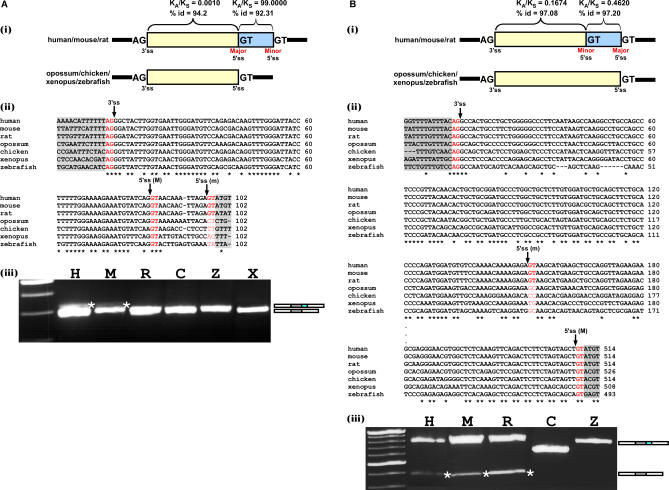
Alternative 3' and 5' splice site (ss) events constitute a significant part of all alternative splicing events. These events were also found to be related to several aberrant splicing diseases. However, only few of the characteristics that distinguish these events from alternative cassette exons are known currently. In this study, we compared the characteristics of constitutive exons, alternative cassette exons, and alternative 3'ss and 5'ss exons. The results revealed that alternative 3'ss and 5'ss exons are an intermediate state between constitutive and alternative cassette exons, where the constitutive side resembles constitutive exons, and the alternative side resembles alternative cassette exons. The results also show that alternative 3'ss and 5'ss exons exhibit low levels of symmetry (frame-preserving), similar to constitutive exons, whereas the sequence between the two alternative splice sites shows high symmetry levels, similar to alternative cassette exons. In addition, flanking intronic conservation analysis revealed that exons whose alternative splice sites are at least nine nucleotides apart show a high conservation level, indicating intronic participation in the regulation of their splicing, whereas exons whose alternative splice sites are fewer than nine nucleotides apart show a low conservation level. Further examination of these exons, spanning seven vertebrate species, suggests an evolutionary model in which the alternative state is a derivative of an ancestral constitutive exon, where a mutation inside the exon or along the flanking intron resulted in the creation of a new splice site that competes with the original one, leading to alternative splice site selection. This model was validated experimentally on four exons, showing that they indeed originated from constitutive exons that acquired a new competing splice site during evolution.
Conflict of interest statement
Figures

References
-
- Consortium TCeS. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science. 1998;282:2012–2018. - PubMed
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921. - PubMed
-
- Graveley BR. Alternative splicing: Increasing diversity in the proteomic world. Trends Genet. 2001;17:100–107. - PubMed
-
- Hastings ML, Krainer AR. Pre-mRNA splicing in the new millennium. Curr Opin Cell Biol. 2001;13:302–309. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
