

Insulin-like growth factor 1 (IGF-1) mediates the effects of enriched environment (EE) on visual cortical development
- PMID: 17534425
- PMCID: PMC1871611
- DOI: 10.1371/journal.pone.0000475
Insulin-like growth factor 1 (IGF-1) mediates the effects of enriched environment (EE) on visual cortical development
Abstract
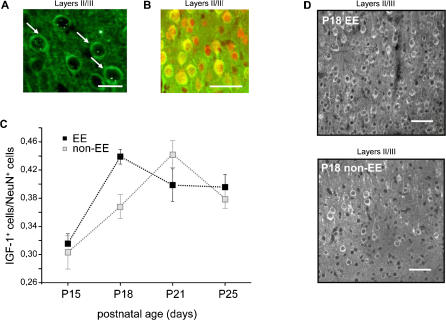
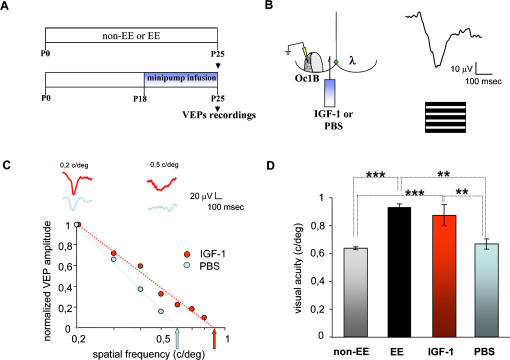
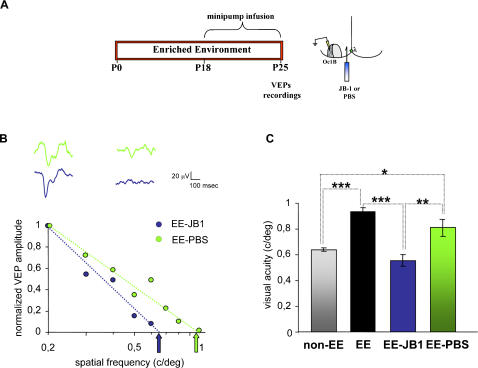
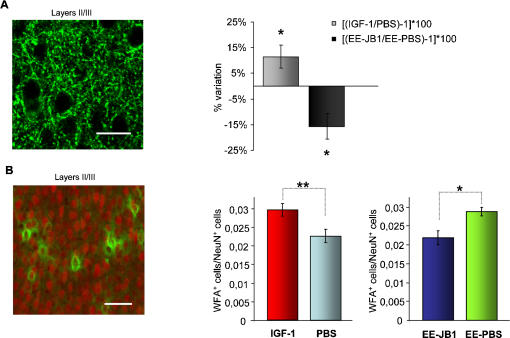
Enriched environment (EE) has been recently shown to affect visual cortex development and plasticity, and to prevent dark rearing effects. The factors mediating EE effects on visual cortical development and plasticity are still unclear. We have investigated whether IGF-1 is involved in mediating EE effects on the developing visual cortex. We show that EE increases the number of IGF-1 positive neurons in the visual cortex at P18. Increasing IGF-1 in the visual cortex of non-EE rats by means of osmotic minipumps implanted at P18 mimics EE effects, accelerating visual acuity development, assessed with Visual Evoked Potentials (VEPs). Blocking IGF-1 action in the visual cortex of EE rats by means of the IGF-1 receptor antagonist JB1 from P18 completely blocks EE action on visual acuity development. These results show that IGF-1 is a key factor mediating EE effects on visual cortical development. We then show that IGF-1 affects GAD65 immunoreactivity in perisomatic innervation and the condensation of Chondroitin Sulphate Proteoglycans (CSPGs) in perineuronal nets (PNNs) in the visual cortex. This suggests that IGF-1 action in mediating EE effects could be exerted through the modulation of intracortical inhibitory circuitry and PNN development.
Conflict of interest statement
Figures
Similar articles
-
Early IGF-1 primes visual cortex maturation and accelerates developmental switch between NKCC1 and KCC2 chloride transporters in enriched animals.Neuropharmacology. 2017 Feb;113(Pt A):167-177. doi: 10.1016/j.neuropharm.2016.02.034. Epub 2016 May 11. Neuropharmacology. 2017. PMID: 26924708
-
Experience-dependent reactivation of ocular dominance plasticity in the adult visual cortex.Exp Neurol. 2010 Nov;226(1):100-9. doi: 10.1016/j.expneurol.2010.08.009. Epub 2010 Aug 14. Exp Neurol. 2010. PMID: 20713044
-
Environmental enrichment potentiates thalamocortical transmission and plasticity in the adult rat visual cortex.J Neurosci Res. 2010 Nov 1;88(14):3048-59. doi: 10.1002/jnr.22461. J Neurosci Res. 2010. PMID: 20722076
-
Expression of perineuronal nets, parvalbumin and protein tyrosine phosphatase σ in the rat visual cortex during development and after BFD.Curr Eye Res. 2013 Oct;38(10):1083-94. doi: 10.3109/02713683.2013.803287. Epub 2013 May 29. Curr Eye Res. 2013. PMID: 23718120
-
Enrich the environment to empower the brain.Trends Neurosci. 2009 Apr;32(4):233-9. doi: 10.1016/j.tins.2008.12.004. Epub 2009 Mar 4. Trends Neurosci. 2009. PMID: 19268375 Review.
Cited by
-
The calm mouse: an animal model of stress reduction.Mol Med. 2012 May 9;18(1):606-17. doi: 10.2119/molmed.2012.00053. Mol Med. 2012. PMID: 22398685 Free PMC article.
-
Effect of maternal care on hearing onset induced by developmental changes in the auditory periphery.J Neurosci. 2014 Mar 26;34(13):4528-33. doi: 10.1523/JNEUROSCI.4188-13.2014. J Neurosci. 2014. PMID: 24671998 Free PMC article.
-
Food restriction enhances visual cortex plasticity in adulthood.Nat Commun. 2011;2:320. doi: 10.1038/ncomms1323. Nat Commun. 2011. PMID: 21587237
-
Plasticity in the adult brain: lessons from the visual system.Exp Brain Res. 2009 Jan;192(3):335-41. doi: 10.1007/s00221-008-1509-3. Epub 2008 Jul 31. Exp Brain Res. 2009. PMID: 18668231 Review.
-
Synaptic and circuit development of the primary sensory cortex.Exp Mol Med. 2018 Apr 9;50(4):1-9. doi: 10.1038/s12276-018-0029-x. Exp Mol Med. 2018. PMID: 29628505 Free PMC article. Review.
References
-
- Rosenzweig MR. Environmental complexity, cerebral change, and behavior. Am Psychol. 1966;21:321–332. - PubMed
-
- Rosenzweig MR, Bennett EL. Psychobiology of plasticity: effects of training and experience on brain and behavior. Behav Brain Res. 1996;78:57–65. - PubMed
-
- Rampon C, Tang YP, Goodhouse J, Shimizu E, Kyin M, et al. Enrichment induces structural changes and recovery from nonspatial memory deficits in CA1 NMDAR1-knockout mice. Nat Neurosci. 2000;3:238–244. - PubMed
-
- van Praag H, Kempermann G, Gage FH. Neural consequences of environmental enrichment. Nat Rev Neurosci. 2000;1:191–198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
