

A database of wing diversity in the Hawaiian Drosophila
- PMID: 17534437
- PMCID: PMC1872047
- DOI: 10.1371/journal.pone.0000487
A database of wing diversity in the Hawaiian Drosophila
Abstract
Background: Within genus Drosophila, the endemic Hawaiian species offer some of the most dramatic examples of morphological and behavioral evolution. The advent of the Drosophila grimshawi genome sequence permits genes of interest to be readily cloned from any of the hundreds of species of Hawaiian Drosophila, offering a powerful comparative approach to defining molecular mechanisms of species evolution. A key step in this process is to survey the Hawaiian flies for characters whose variation can be associated with specific candidate genes. The wings provide an attractive target for such studies: Wings are essentially two dimensional, and genes controlling wing shape, vein specification, pigment production, and pigment pattern evolution have all been identified in Drosophila.
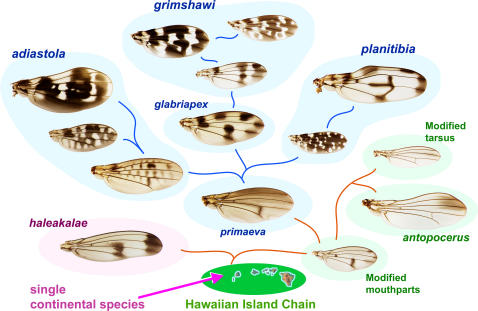
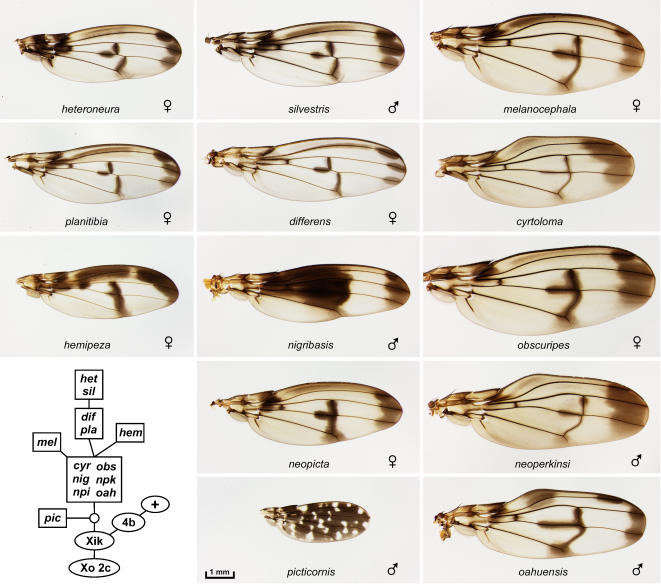
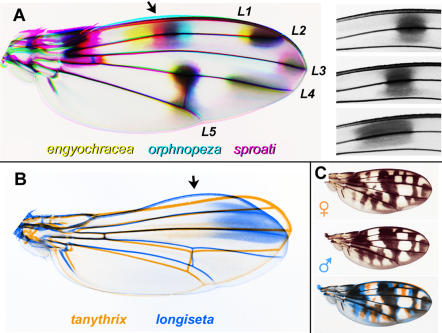
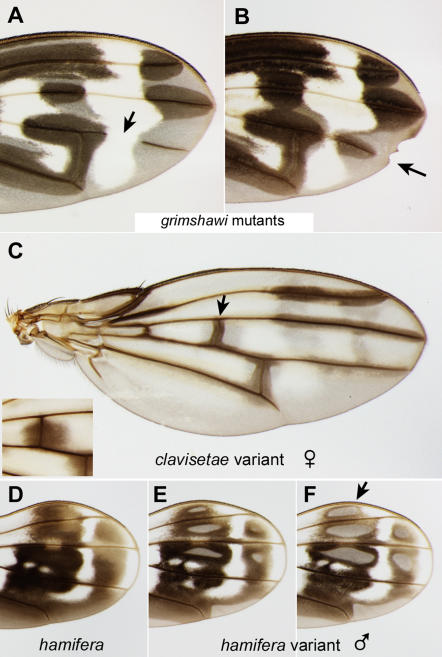
Methodology/principal findings: We present a photographic database of over 180 mounted, adult wings from 73 species of Hawaiian Drosophila. The image collection, available at FlyBase.org, includes 53 of the 112 known species of "picture wing" Drosophila, and several species from each of the other major Hawaiian groups, including the modified mouthparts, modified tarsus, antopocerus, and haleakalae (fungus feeder) groups. Direct image comparisons show that major wing shape changes can occur even between closely related species, and that pigment pattern elements can vary independently of each other. Among the 30 species closest to grimshawi, diverse visual effects are achieved by altering a basic pattern of seven wing spots. Finally, we document major pattern variations within species, which appear to result from reduced diffusion of pigment precursors through the wing blade.
Conclusions/significance: The database highlights the striking variation in size, shape, venation, and pigmentation in Hawaiian Drosophila, despite their generally low levels of DNA sequence divergence. In several independent lineages, highly complex patterns are derived from simple ones. These lineages offer a promising model system to study the evolution of complexity.
Conflict of interest statement
Figures





Similar articles
-
Ultrastructural variation and adaptive evolution of the ovipositor in the endemic Hawaiian Drosophilidae.J Morphol. 2018 Dec;279(12):1725-1752. doi: 10.1002/jmor.20884. Epub 2018 Nov 5. J Morphol. 2018. PMID: 30397938
-
Automated measurement of Drosophila wings.BMC Evol Biol. 2003 Dec 11;3:25. doi: 10.1186/1471-2148-3-25. BMC Evol Biol. 2003. PMID: 14670094 Free PMC article.
-
Hawaiian Drosophila genomes: size variation and evolutionary expansions.Genetica. 2016 Feb;144(1):107-24. doi: 10.1007/s10709-016-9882-5. Epub 2016 Jan 20. Genetica. 2016. PMID: 26790663
-
Evolution of wing pigmentation in Drosophila: Diversity, physiological regulation, and cis-regulatory evolution.Dev Growth Differ. 2020 Jun;62(5):269-278. doi: 10.1111/dgd.12661. Epub 2020 Apr 4. Dev Growth Differ. 2020. PMID: 32171022 Free PMC article. Review.
-
Developmental basis for vein pattern variations in insect wings.Int J Dev Biol. 2003;47(7-8):653-63. Int J Dev Biol. 2003. PMID: 14756341 Review.
Cited by
-
Microsatellite repeat instability fuels evolution of embryonic enhancers in Hawaiian Drosophila.PLoS One. 2014 Jun 30;9(6):e101177. doi: 10.1371/journal.pone.0101177. eCollection 2014. PLoS One. 2014. PMID: 24978198 Free PMC article.
-
DrosoPhyla: Resources for Drosophilid Phylogeny and Systematics.Genome Biol Evol. 2021 Aug 3;13(8):evab179. doi: 10.1093/gbe/evab179. Genome Biol Evol. 2021. PMID: 34343293 Free PMC article.
-
Sequencing degraded DNA from non-destructively sampled museum specimens for RAD-tagging and low-coverage shotgun phylogenetics.PLoS One. 2014 May 14;9(5):e96793. doi: 10.1371/journal.pone.0096793. eCollection 2014. PLoS One. 2014. PMID: 24828244 Free PMC article.
-
Inverse resource allocation between vision and olfaction across the genus Drosophila.Nat Commun. 2019 Mar 11;10(1):1162. doi: 10.1038/s41467-019-09087-z. Nat Commun. 2019. PMID: 30858374 Free PMC article.
-
Stable structural color patterns displayed on transparent insect wings.Proc Natl Acad Sci U S A. 2011 Jan 11;108(2):668-73. doi: 10.1073/pnas.1017393108. Epub 2011 Jan 3. Proc Natl Acad Sci U S A. 2011. PMID: 21199954 Free PMC article.
References
-
- Carson HL, Kaneshiro KY. Drosophila of Hawaii: Systematics and ecological genetics. Ann Rev Ecology Systematics. 1976;7:311–345.
-
- Carson HL, Clague DA. Geology and biogeography of the Hawaiian Islands. In: Wagner WL, Funk VA, editors. Hawaiian Biogeography: Evolution on a hot spot archipelago. Washington D.C.: Smithsonian Institution Press; 1995. pp. 14–29.
-
- Carson HL. Sexual selection: A driver of genetic change in Hawaiian Drosophila. J Hered. 1997;88:343–352. - PubMed
-
- Remsen J, DeSalle R. Character congruence of multiple data partitions and the origin of the Hawaiian Drosophilidae. Mol Phylogenet Evol. 1998;9:225–235. - PubMed
-
- Craddock EM. Speciation processes in the adaptive radiation of Hawaiian plants and animals. Evol Biol. 2000;31:1–43.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
