

Stepwise proteolysis liberates tau fragments that nucleate the Alzheimer-like aggregation of full-length tau in a neuronal cell model
- PMID: 17535890
- PMCID: PMC1891218
- DOI: 10.1073/pnas.0703676104
Stepwise proteolysis liberates tau fragments that nucleate the Alzheimer-like aggregation of full-length tau in a neuronal cell model
Abstract
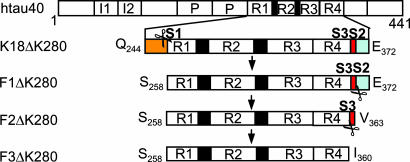
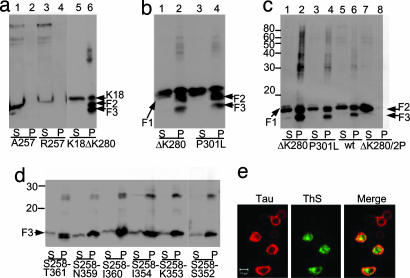
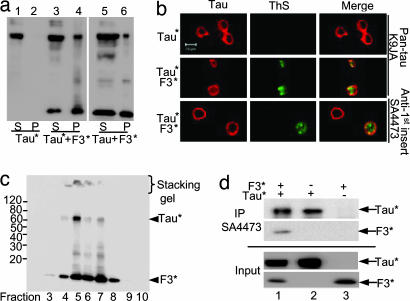
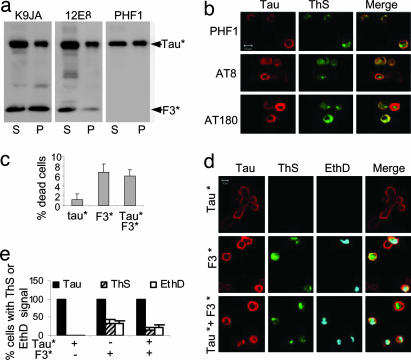
Tau is a highly soluble protein, yet it aggregates abnormally in Alzheimer's disease. Here, we address the question of proteolytic processing of tau and the nucleation of aggregates by tau fragments. We show in neuronal cell models that fragments of the repeat domain of tau containing mutations of FTDP17 (frontotemporal dementia with parkinsonism linked to chromosome 17), produced by endogenous proteases, can induce the aggregation of full-length tau. Fragments are generated by successive cleavages, first N-terminally between K257 and S258, then C-terminally around residues 353-364; conversely, when the N-terminal cleavage is inhibited, no fragmentation and aggregation takes place. The C-terminal truncation and the coaggregation of fragments with full-length tau depends on the propensity for beta-structure. The aggregation is modulated by phosphorylation but does not depend on it. Aggregation but not fragmentation as such is toxic to cells; conversely, toxicity can be prevented by inhibiting either aggregation or proteolysis. The results reveal a novel pathway of abnormal tau aggregation in neuronal cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Lee VM, Goedert M, Trojanowski JQ. Annu Rev Neurosci. 2001;24:1121–1159. - PubMed
-
- Braak H, Braak E. Acta Neuropathol (Berl) 1991;82:239–259. - PubMed
-
- Spillantini MG, Goedert M. Trends Neurosci. 1998;21:428–433. - PubMed
-
- Lee VM, Trojanowski JQ. J Alzheimer's Dis. 2006;9:257–262. - PubMed
-
- LaFerla FM, Oddo S. Trends Mol Med. 2005;11:170–176. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
