

Receptor-mediated activation of nitric oxide synthesis by arginine in endothelial cells
- PMID: 17535904
- PMCID: PMC1891228
- DOI: 10.1073/pnas.0506824104
Receptor-mediated activation of nitric oxide synthesis by arginine in endothelial cells
Abstract
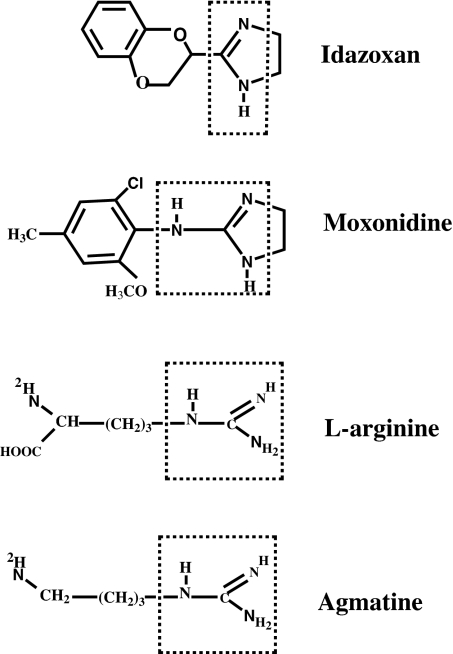
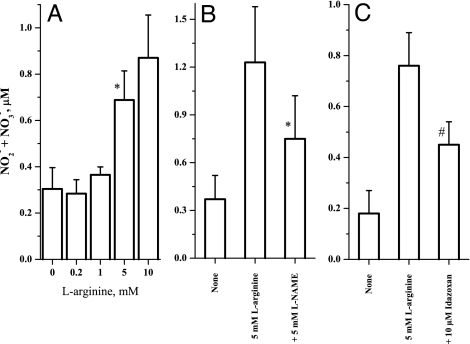
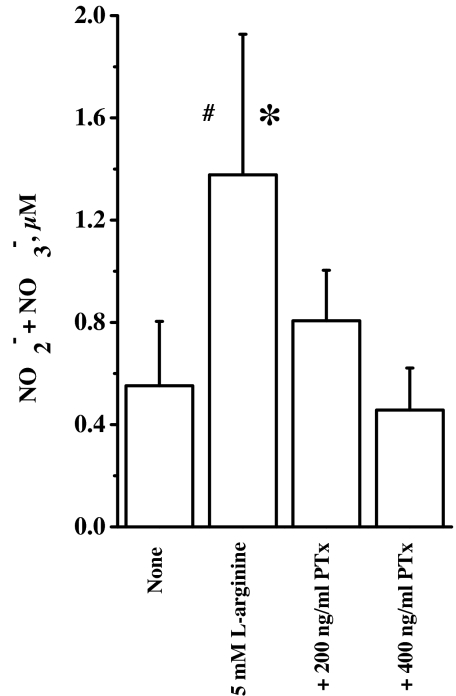
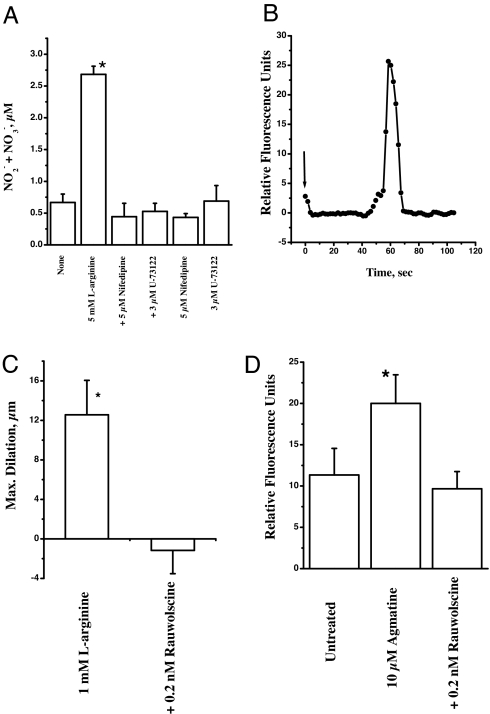
Arginine contains the guanidinium group and thus has structural similarity to ligands of imidazoline and alpha-2 adrenoceptors (alpha-2 AR). Therefore, we investigated the possibility that exogenous arginine may act as a ligand for these receptors in human umbilical vein endothelial cells and activate intracellular nitric oxide (NO) synthesis. Idazoxan, a mixed antagonist of imidazoline and alpha-2 adrenoceptors, partly inhibited L-arginine-initiated NO formation as measured by a Griess reaction. Rauwolscine, a highly specific antagonist of alpha-2 AR, at very low concentrations completely inhibited NO formation. Like L-arginine, agmatine (decarboxylated arginine) also activated NO synthesis, however, at much lower concentrations. We found that dexmedetomidine, a specific agonist of alpha-2 AR was very potent in activating cellular NO, thus indicating a possible role for alpha-2 AR in L-arginine-mediated NO synthesis. D-arginine also activated NO production and could be inhibited by imidazoline and alpha-2 AR antagonists, thus indicating nonsubstrate actions of arginine. Pertussis toxin, an inhibitor of G proteins, attenuated L-arginine-mediated NO synthesis, thus indicating mediation via G proteins. L-type Ca(2+) channel blocker nifedipine and phospholipase C inhibitor U73122 inhibited NO formation and thus implicated participation of a second messenger pathway. Finally, in isolated rat gracilis vessels, rauwolscine completely inhibited the L-arginine-initiated vessel relaxation. Taken together, these data provide evidence for binding of arginine to membrane receptor(s), leading to the activation of endothelial NO synthase (eNOS) NO production through a second messenger pathway. These findings provide a previously unrecognized mechanistic explanation for the beneficial effects of L-arginine in the cardiovascular system and thus provide new potential avenues for therapeutic development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Mechanisms underlying beta2-adrenoceptor-mediated nitric oxide generation by human umbilical vein endothelial cells.J Physiol. 2006 Oct 15;576(Pt 2):585-94. doi: 10.1113/jphysiol.2006.115998. Epub 2006 Jul 27. J Physiol. 2006. PMID: 16873402 Free PMC article.
-
Activation of nitric oxide synthase by beta 2-adrenoceptors in human umbilical vein endothelium in vitro.Br J Pharmacol. 1999 Apr;126(8):1872-80. doi: 10.1038/sj.bjp.0702512. Br J Pharmacol. 1999. PMID: 10372832 Free PMC article.
-
Alpha-2 adrenoceptors and imidazoline receptors in cardiomyocytes mediate counterbalancing effect of agmatine on NO synthesis and intracellular calcium handling.J Mol Cell Cardiol. 2014 Mar;68:66-74. doi: 10.1016/j.yjmcc.2013.12.030. Epub 2014 Jan 9. J Mol Cell Cardiol. 2014. PMID: 24412533
-
[Molecular targets for searching of endothelial-protective substances].Bioorg Khim. 2014 Sep-Oct;40(5):515-27. doi: 10.1134/s1068162014050069. Bioorg Khim. 2014. PMID: 25895347 Review. Russian.
-
Cardiovascular effects of L-arginine.Curr Opin Nephrol Hypertens. 1998 Jan;7(1):63-70. doi: 10.1097/00041552-199801000-00011. Curr Opin Nephrol Hypertens. 1998. PMID: 9442365 Review.
Cited by
-
Decoupling Phase Separation and Fibrillization Preserves Activity of Biomolecular Condensates.Res Sq [Preprint]. 2025 Apr 29:rs.3.rs-6405673. doi: 10.21203/rs.3.rs-6405673/v1. Res Sq. 2025. PMID: 40343340 Free PMC article. Preprint.
-
Characterization and inactivation of an agmatine deiminase from Helicobacter pylori.Bioorg Chem. 2010 Apr;38(2):62-73. doi: 10.1016/j.bioorg.2009.11.004. Epub 2009 Nov 29. Bioorg Chem. 2010. PMID: 20036411 Free PMC article.
-
Interactions of nitric oxide with α2 -adrenoceptors within the locus coeruleus underlie the facilitation of inhibitory avoidance memory by agmatine.Br J Pharmacol. 2016 Sep;173(17):2589-99. doi: 10.1111/bph.13531. Epub 2016 Aug 1. Br J Pharmacol. 2016. PMID: 27273730 Free PMC article.
-
Intracellular α(2C)-adrenoceptors: storage depot, stunted development or signaling domain?Biochim Biophys Acta. 2011 Aug;1813(8):1495-503. doi: 10.1016/j.bbamcr.2011.05.006. Epub 2011 May 14. Biochim Biophys Acta. 2011. PMID: 21605601 Free PMC article. Review.
-
Tetrahydrobiopterin, L-arginine and vitamin C actsynergistically to decrease oxidative stress, increase nitricoxide and improve blood flow after induction of hindlimbischemia in the rat.Mol Med. 2012 May 9;18(1):676-84. doi: 10.2119/molmed.2011.00103. Mol Med. 2012. PMID: 22371305 Free PMC article.
References
-
- Forstermann U, Closs EI, Pollock JS, Nakane M, Schwarz P, Gath I, Kleinert H. Hypertension. 1994;23:1121–1131. - PubMed
-
- Tsikas D, Boger RH, Sandmann J, Bode-Boger SM, Frolich JC. FEBS Lett. 2000;478:1–3. - PubMed
-
- Garcia-Cardena G, Martasek P, Masters BS, Skidd PM, Couet J, Li S, Lisanti MP, Sessa WC. J Biol Chem. 1997;272:25437–25440. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
