

Epigenetic modulation of seizure-induced neurogenesis and cognitive decline
- PMID: 17537967
- PMCID: PMC6672253
- DOI: 10.1523/JNEUROSCI.0110-07.2007
Epigenetic modulation of seizure-induced neurogenesis and cognitive decline
Abstract
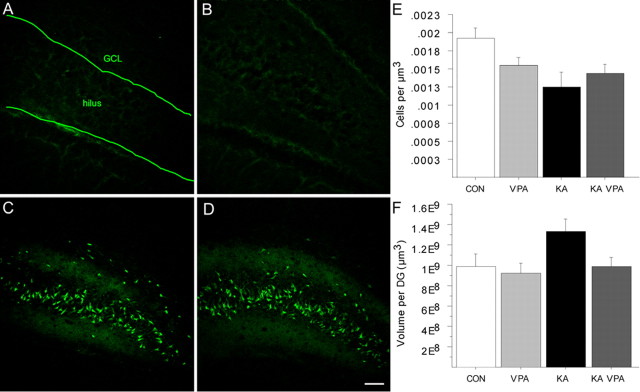
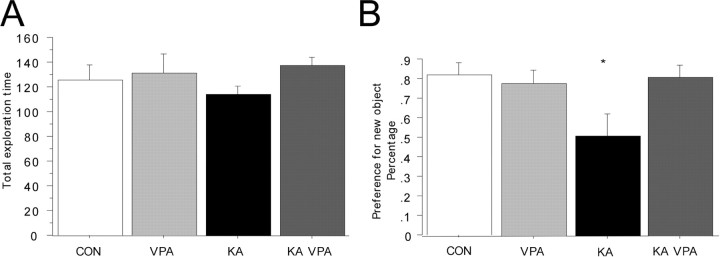
The conceptual understanding of hippocampal function has been challenged recently by the finding that new granule cells are born throughout life in the mammalian dentate gyrus (DG). The number of newborn neurons is dynamically regulated by a variety of factors. Kainic acid-induced seizures, a rodent model of human temporal lobe epilepsy, strongly induce the proliferation of DG neurogenic progenitor cells and are also associated with long-term cognitive impairment. We show here that the antiepileptic drug valproic acid (VPA) potently blocked seizure-induced neurogenesis, an effect that appeared to be mainly mediated by inhibiting histone deacetylases (HDAC) and normalizing HDAC-dependent gene expression within the epileptic dentate area. Strikingly, the inhibition of aberrant neurogenesis protected the animals from seizure-induced cognitive impairment in a hippocampus-dependent learning task. We propose that seizure-generated granule cells have the potential to interfere with hippocampal function and contribute to cognitive impairment caused by epileptic activity within the hippocampal circuitry. Furthermore, our data indicate that the effectiveness of VPA as an antiepileptic drug may be partially explained by the HDAC-dependent inhibition of aberrant neurogenesis induced by seizure activity within the adult hippocampus.
Figures
References
-
- Ballas N, Battaglioli E, Atouf F, Andres ME, Chenoweth J, Anderson ME, Burger C, Moniwa M, Davie JR, Bowers WJ, Federoff HJ, Rose DW, Rosenfeld MG, Brehm P, Mandel G. Regulation of neuronal traits by a novel transcriptional complex. Neuron. 2001;31:353–365. - PubMed
-
- Ben-Ari Y, Cossart R. Kainate, a double agent that generates seizures: two decades of progress. Trends Neurosci. 2000;23:580–587. - PubMed
-
- Bengzon J, Mohapel P, Ekdahl CT, Lindvall O. Neuronal apoptosis after brief and prolonged seizures. Prog Brain Res. 2002;135:111–119. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical