

Rab11A controls the biogenesis of Birbeck granules by regulating Langerin recycling and stability
- PMID: 17538027
- PMCID: PMC1949377
- DOI: 10.1091/mbc.e06-09-0779
Rab11A controls the biogenesis of Birbeck granules by regulating Langerin recycling and stability
Abstract
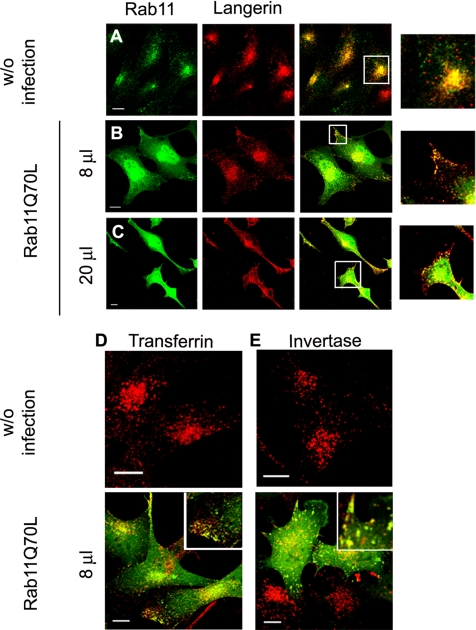
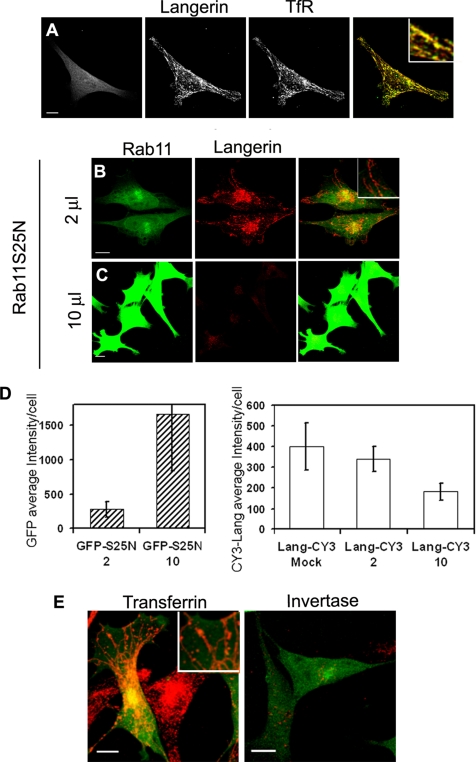
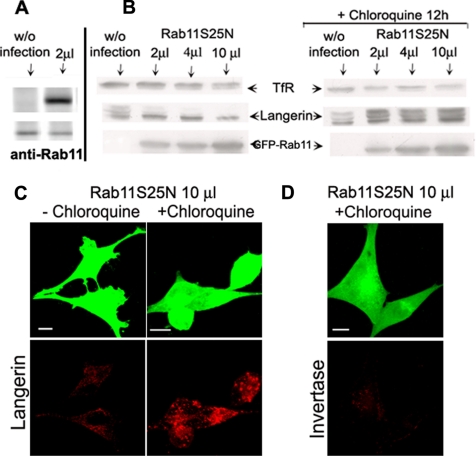
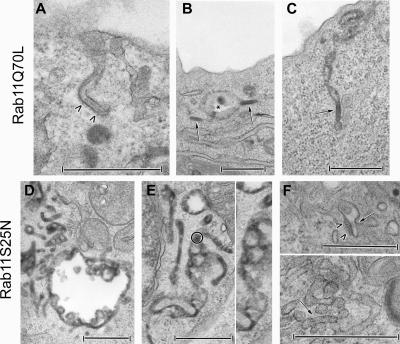
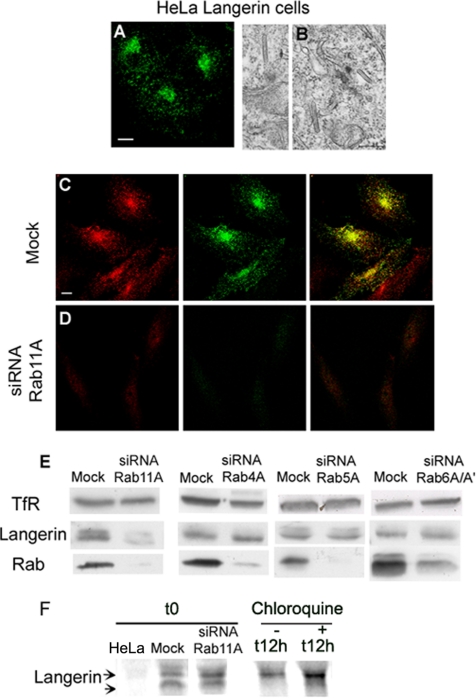
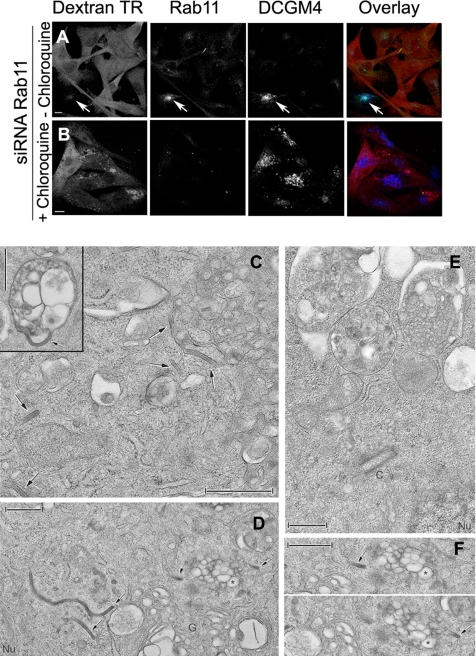
The extent to which Rab GTPases, Rab-interacting proteins, and cargo molecules cooperate in the dynamic organization of membrane architecture remains to be clarified. Langerin, a recycling protein accumulating in the Rab11-positive compartments of Langerhans cells, induces the formation of Birbeck granules (BGs), which are membrane subdomains of the endosomal recycling network. We investigated the role of Rab11A and two members of the Rab11 family of interacting proteins, Rip11 and RCP, in Langerin traffic and the biogenesis of BGs. The overexpression of a dominant-negative Rab11A mutant or Rab11A depletion strongly influenced Langerin traffic and stability and the formation of BGs, whereas modulation of other Rab proteins involved in dynamic regulation of the endocytic-recycling pathway had no effect. Impairment of Rab11A function led to a missorting of Langerin to lysosomal compartments, but inhibition of Langerin degradation by chloroquine did not restore the formation of BGs. Loss of RCP, but not of Rip11, also had a modest, but reproducible effect on Langerin stability and BG biogenesis, pointing to a role for Rab11A-RCP complexes in these events. Our results show that Rab11A and Langerin are required for BG biogenesis, and they illustrate the role played by a Rab GTPase in the formation of a specialized subcompartment within the endocytic-recycling system.
Figures

References
-
- Agard D. A., Hiraoka Y., Shaw P., Sedat J. Fluorescence microscopy in three dimensions. Methods Cell Biol. 1989;30:353–377. - PubMed
-
- Angenieux C., et al. The cellular pathway of CD1e in immature and maturing dendritic cells. Traffic. 2005;6:286–302. - PubMed
-
- Birbeck M. S., Breathnach A. S., Everall J. D. An electron microscope study of basal melanocytes and high-level clear cells (Langerhans cells) in vitiligo. J. Invest. Dermatol. 1961;37:51–63.
-
- Calhoun B. C., Lapierre L. A., Chew C. S., Goldenring J. R. Rab11a redistributes to apical secretory canaliculus during stimulation of gastric parietal cells. Am. J. Physiol. 1998;275:C163–C170. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
