

Biomechanics and biophysics of cancer cells
- PMID: 17540628
- PMCID: PMC2917191
- DOI: 10.1016/j.actbio.2007.04.002
Biomechanics and biophysics of cancer cells
Abstract
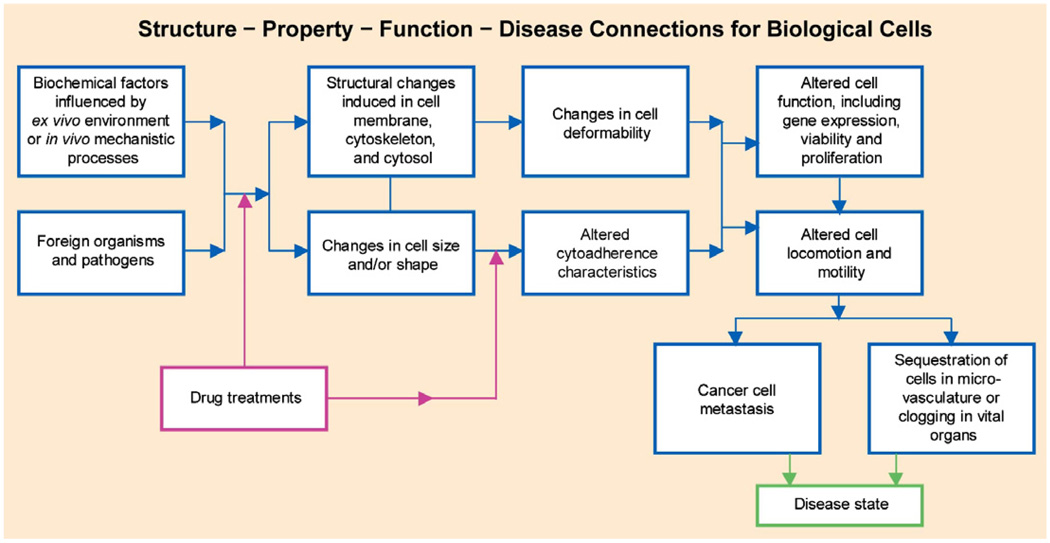
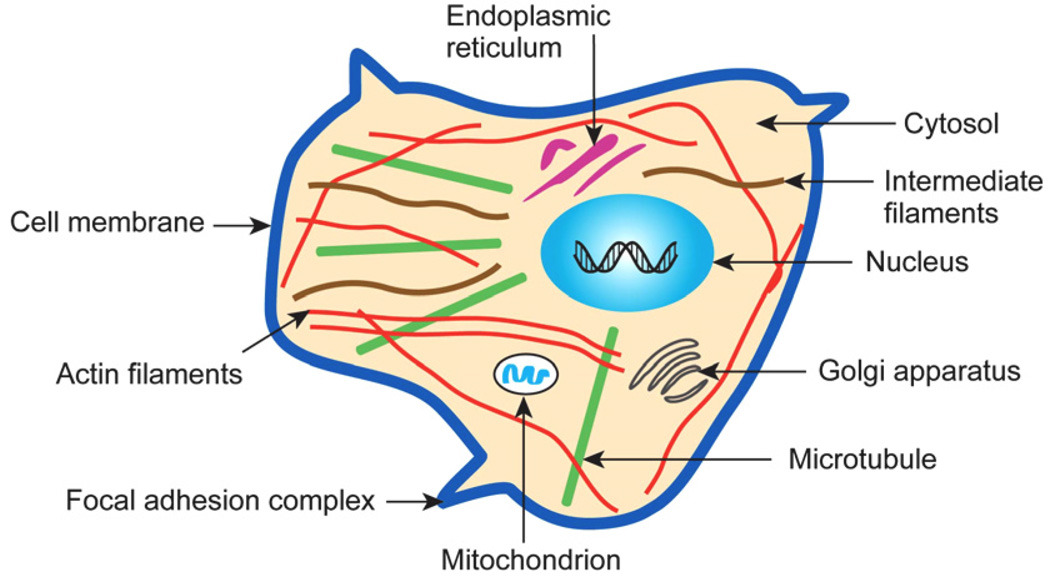
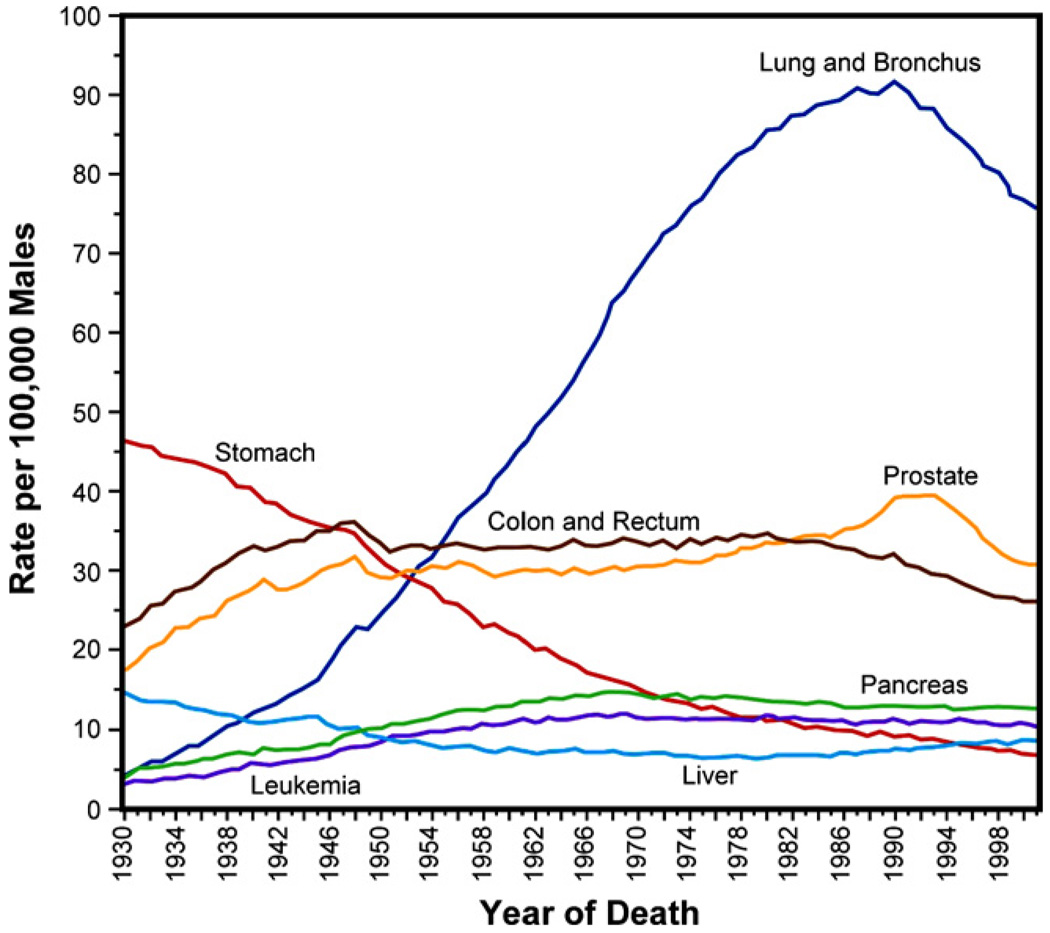
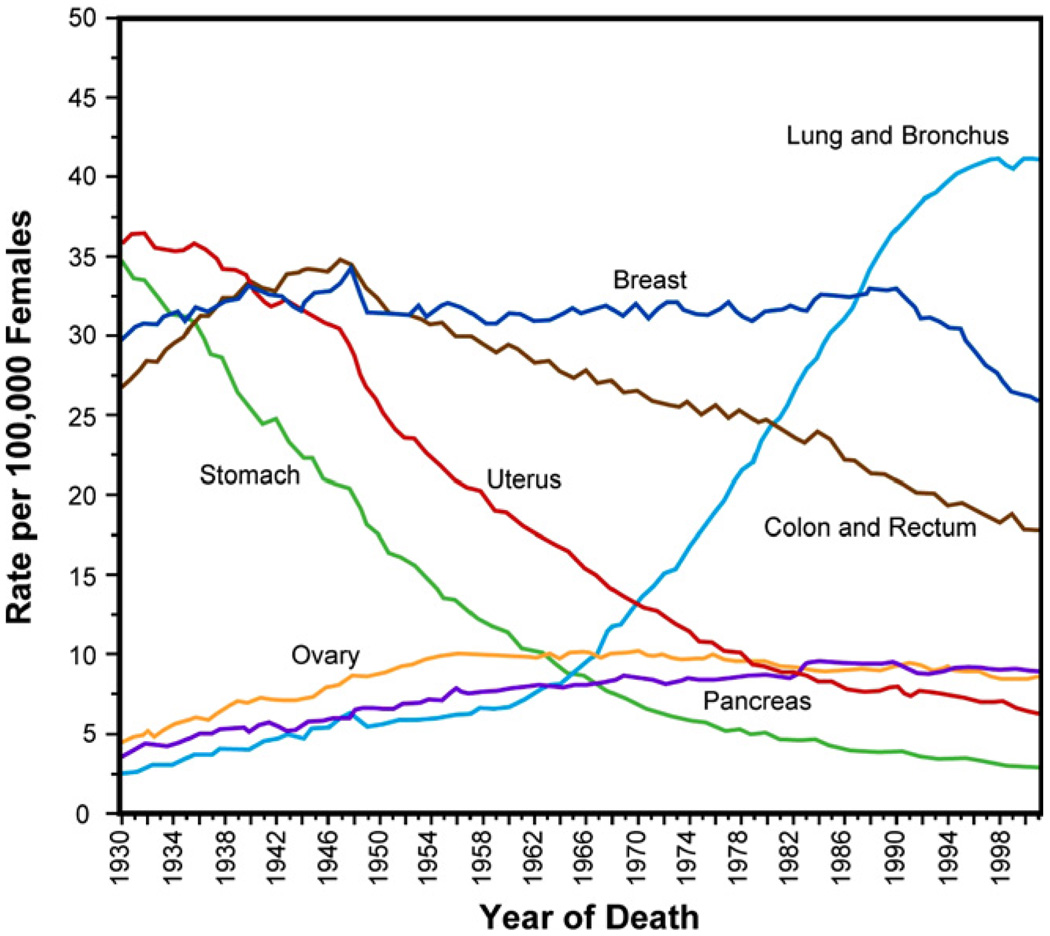
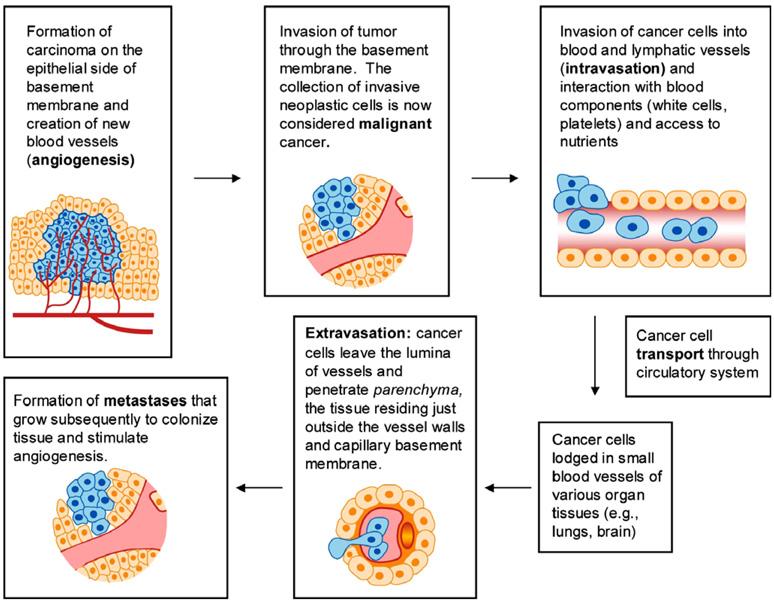
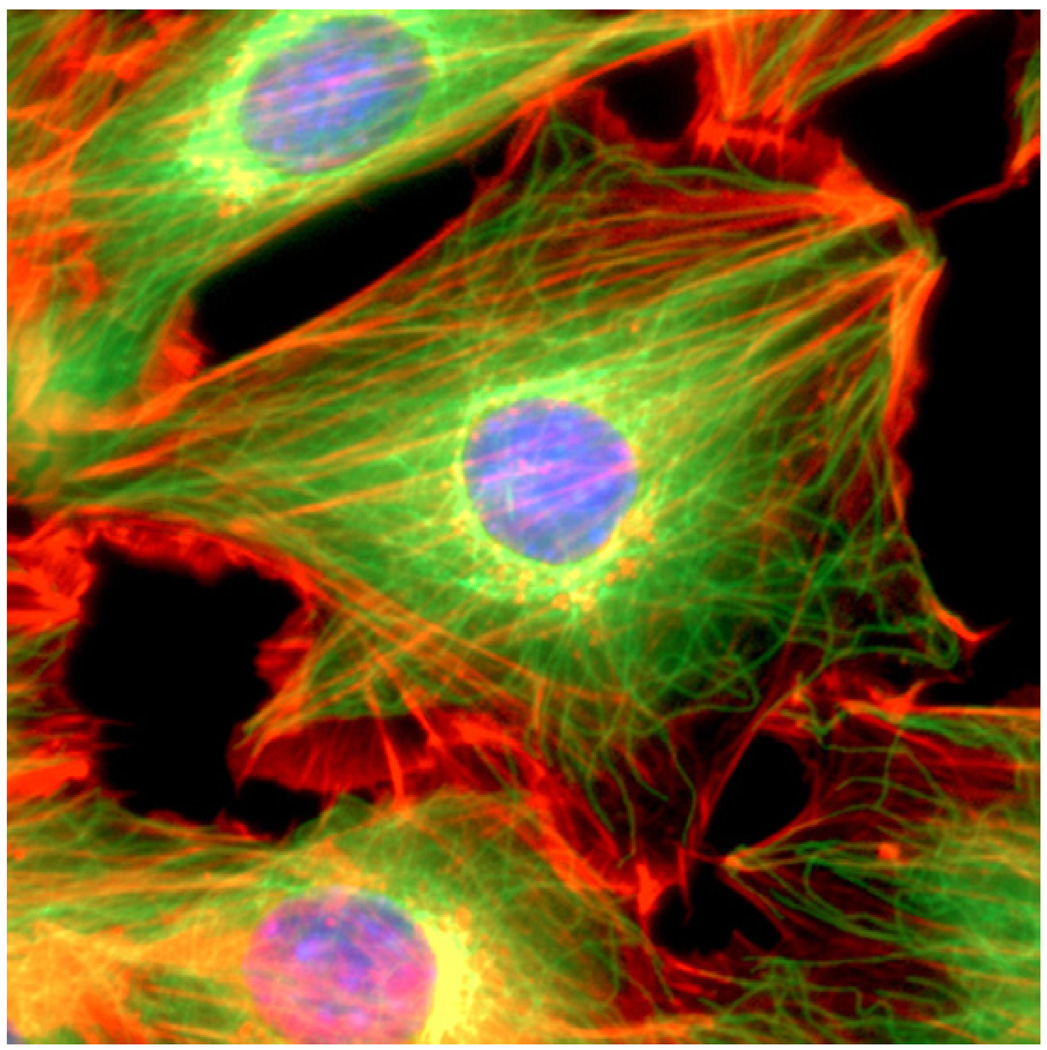
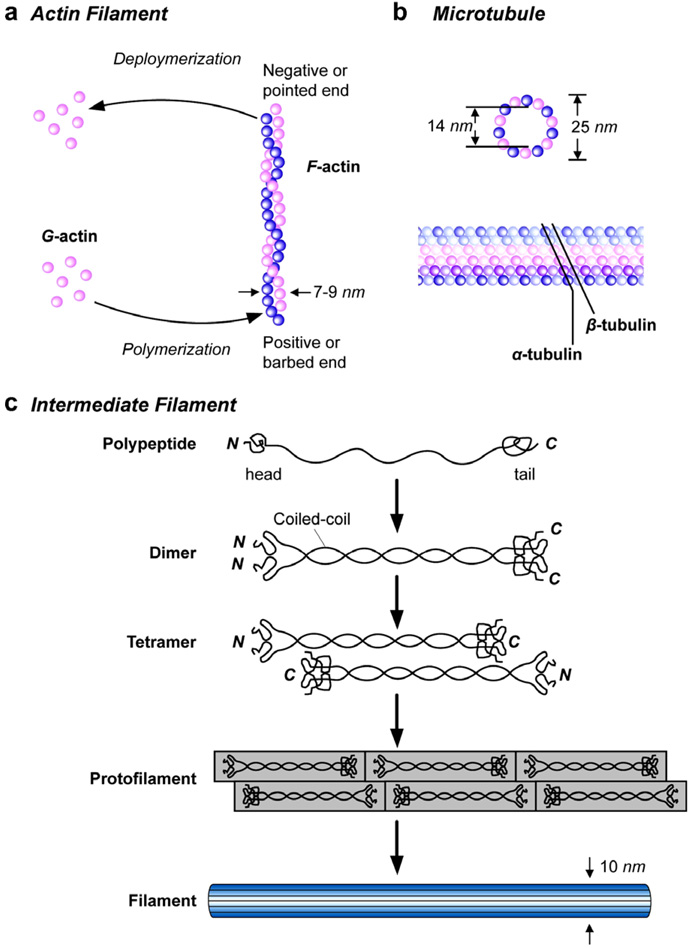
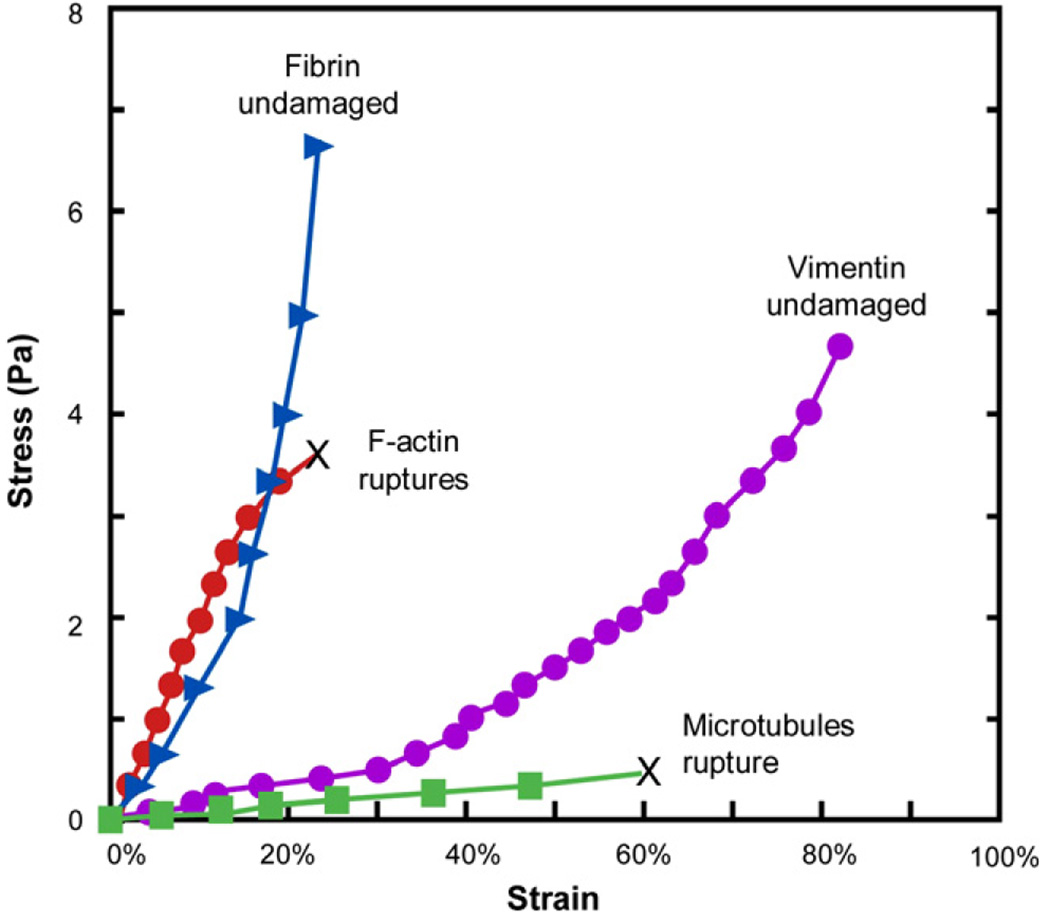
The past decade has seen substantial growth in research into how changes in the biomechanical and biophysical properties of cells and subcellular structures influence, and are influenced by, the onset and progression of human diseases. This paper presents an overview of the rapidly expanding, nascent field of research that deals with the biomechanics and biophysics of cancer cells. The review begins with some key observations on the biology of cancer cells and on the role of actin microfilaments, intermediate filaments and microtubule biopolymer cytoskeletal components in influencing cell mechanics, locomotion, differentiation and neoplastic transformation. In order to set the scene for mechanistic discussions of the connections among alterations to subcellular structures, attendant changes in cell deformability, cytoadherence, migration, invasion and tumor metastasis, a survey is presented of the various quantitative mechanical and physical assays to extract the elastic and viscoelastic deformability of cancer cells. Results available in the literature on cell mechanics for different types of cancer are then reviewed. Representative case studies are presented next to illustrate how chemically induced cytoskeletal changes, biomechanical responses and signals from the intracellular regions act in concert with the chemomechanical environment of the extracellular matrix and the molecular tumorigenic signaling pathways to effect malignant transformations. Results are presented to illustrate how changes to cytoskeletal architecture induced by cancer drugs and chemotherapy regimens can significantly influence cell mechanics and disease state. It is reasoned through experimental evidence that greater understanding of the mechanics of cancer cell deformability and its interactions with the extracellular physical, chemical and biological environments offers enormous potential for significant new developments in disease diagnostics, prophylactics, therapeutics and drug efficacy assays.
Figures
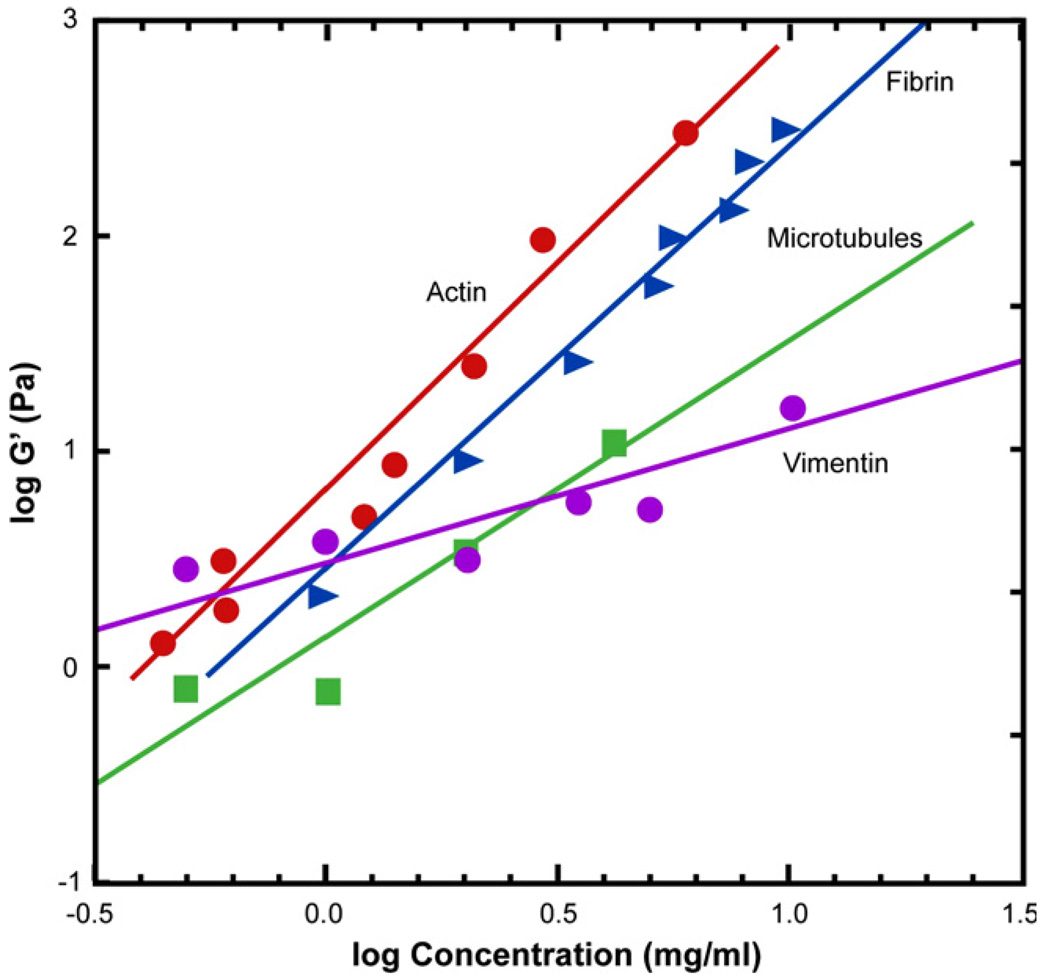
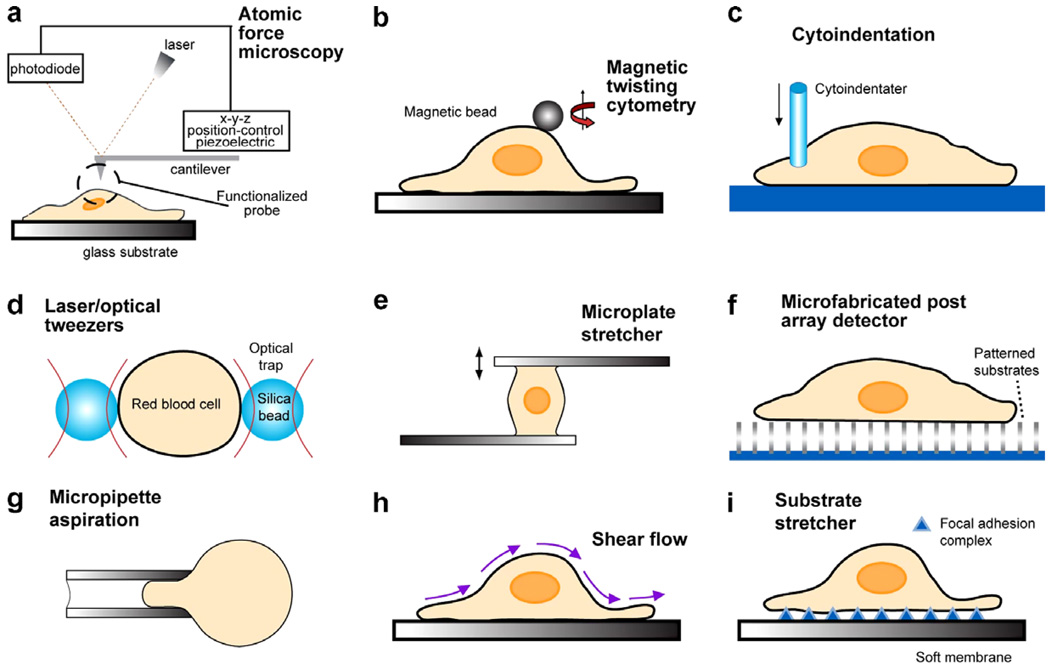
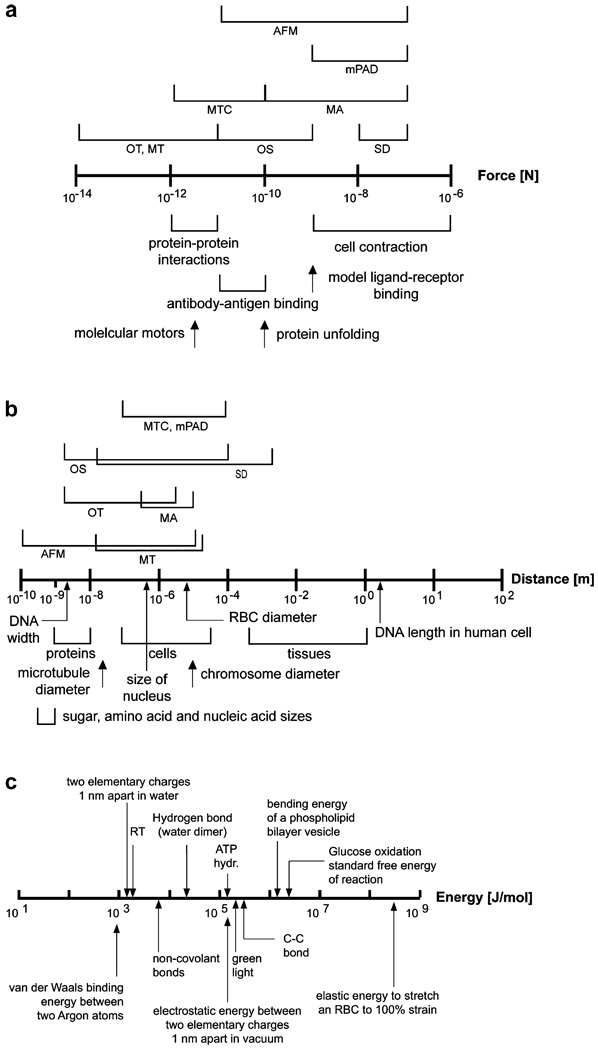
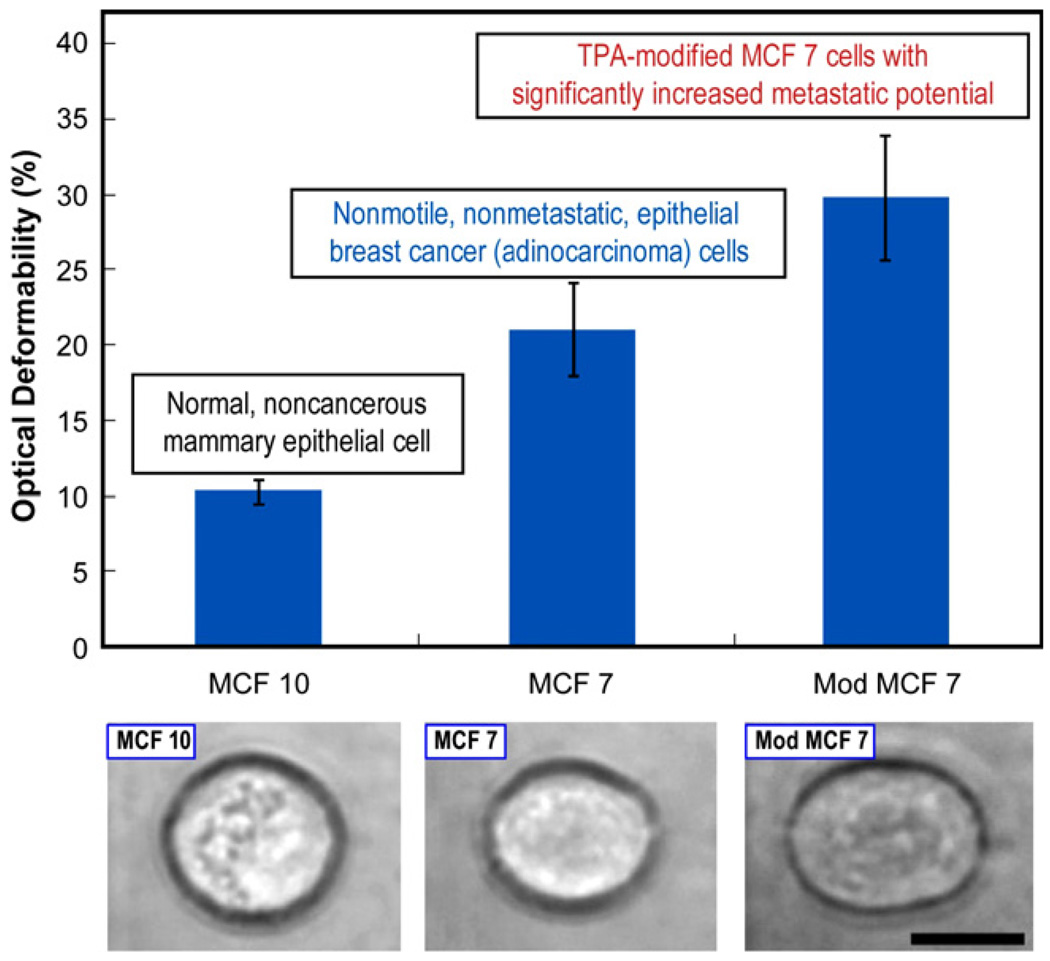
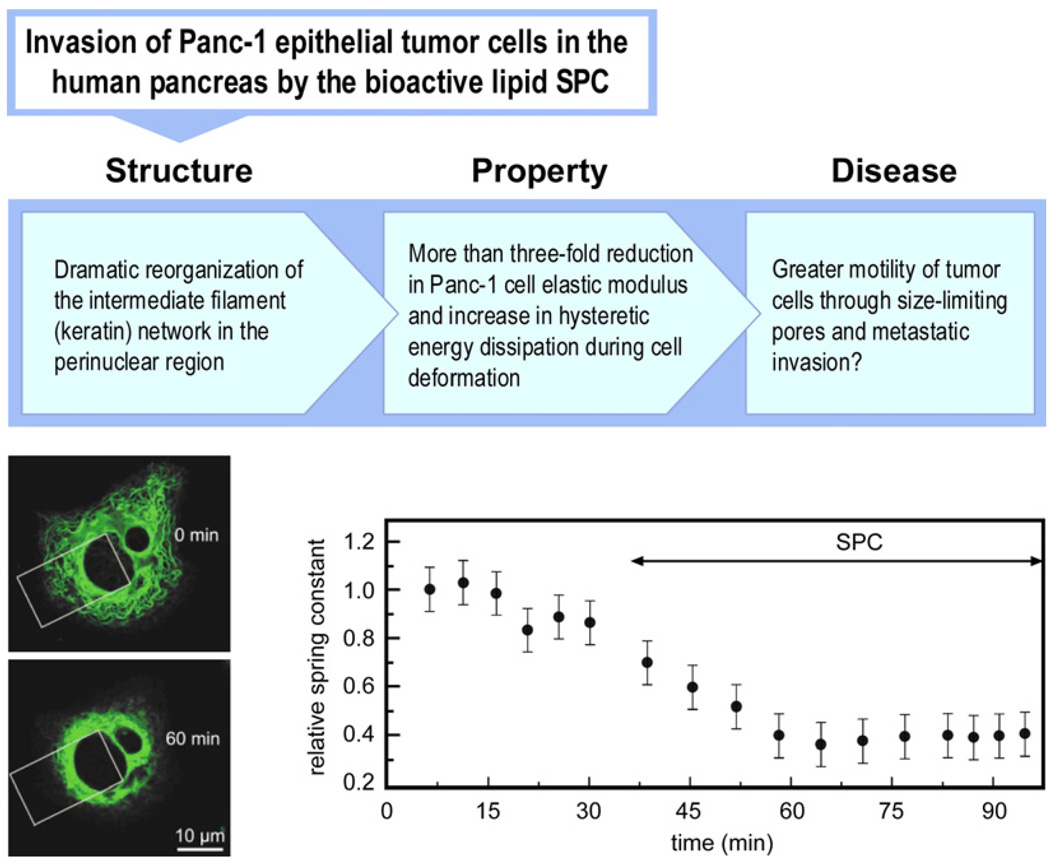
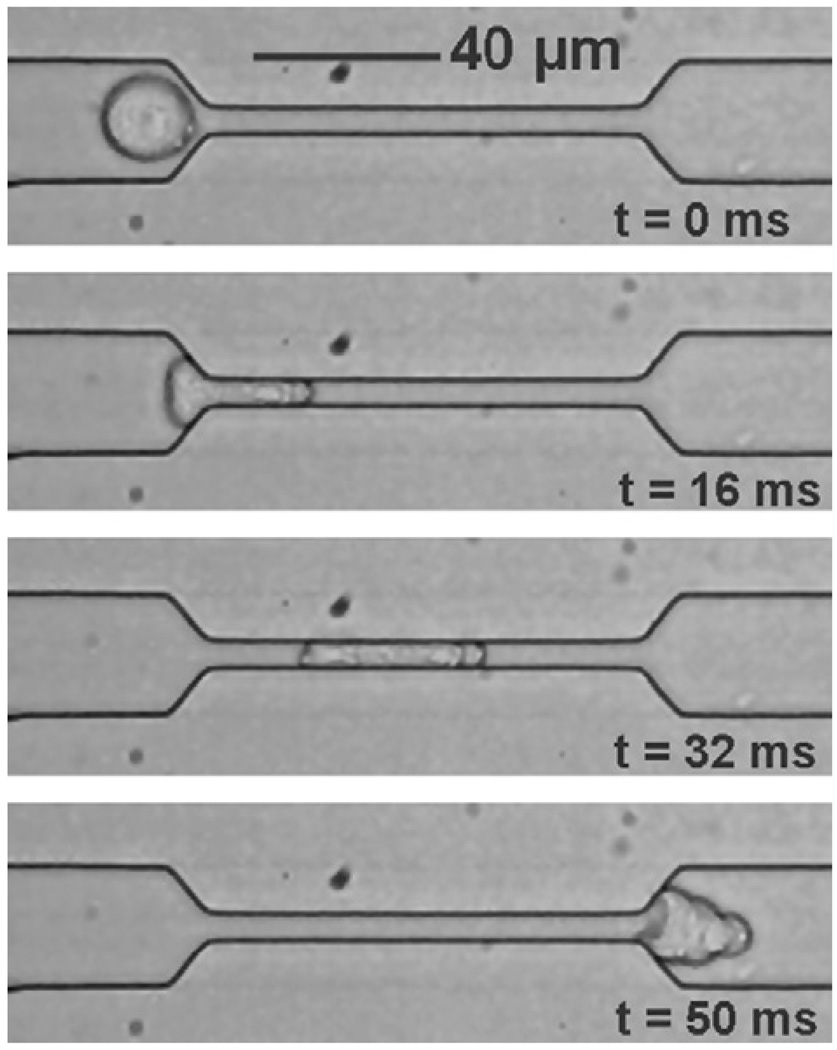
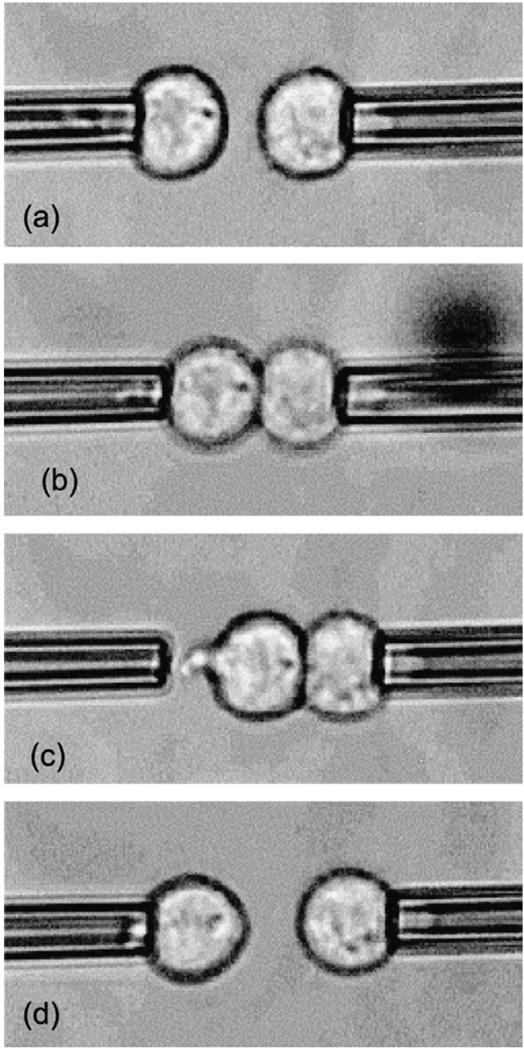
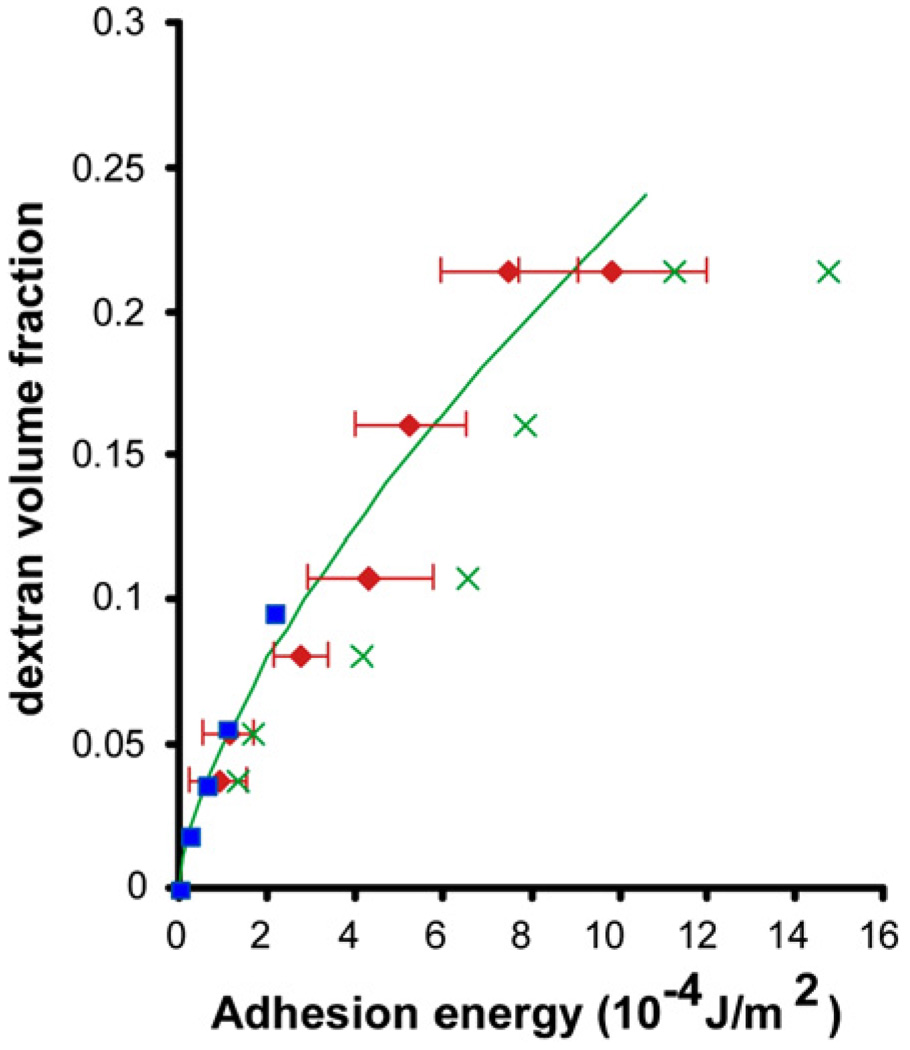
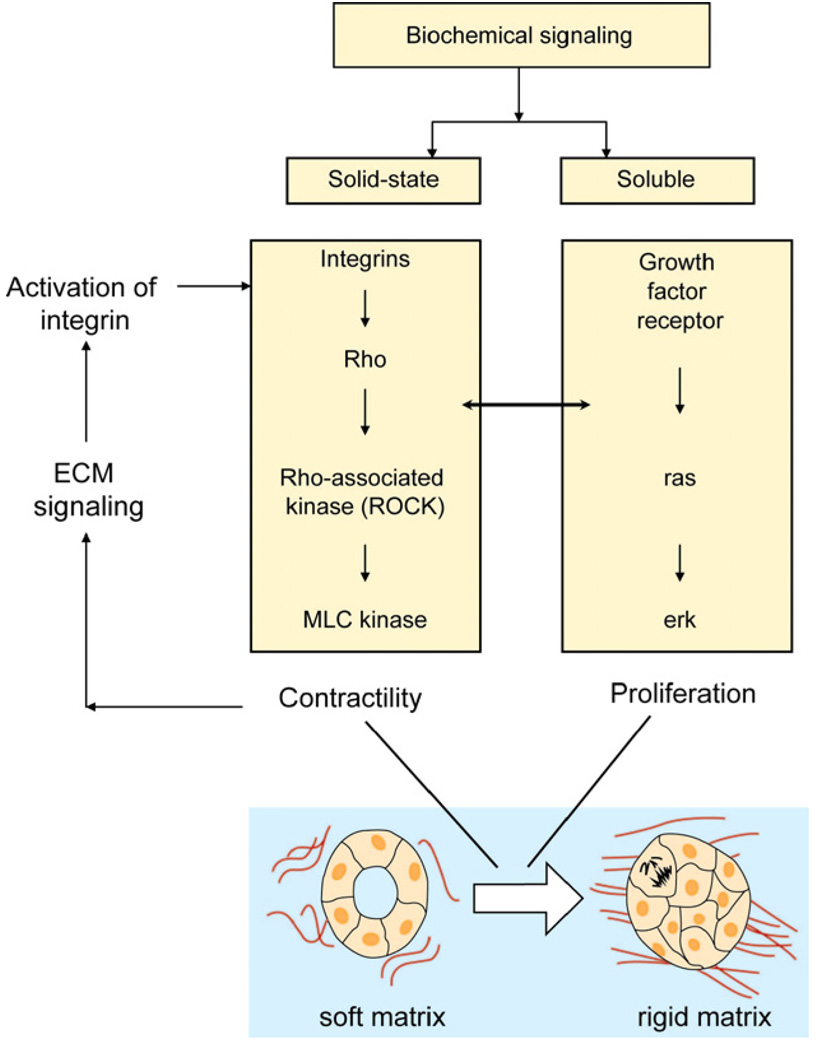
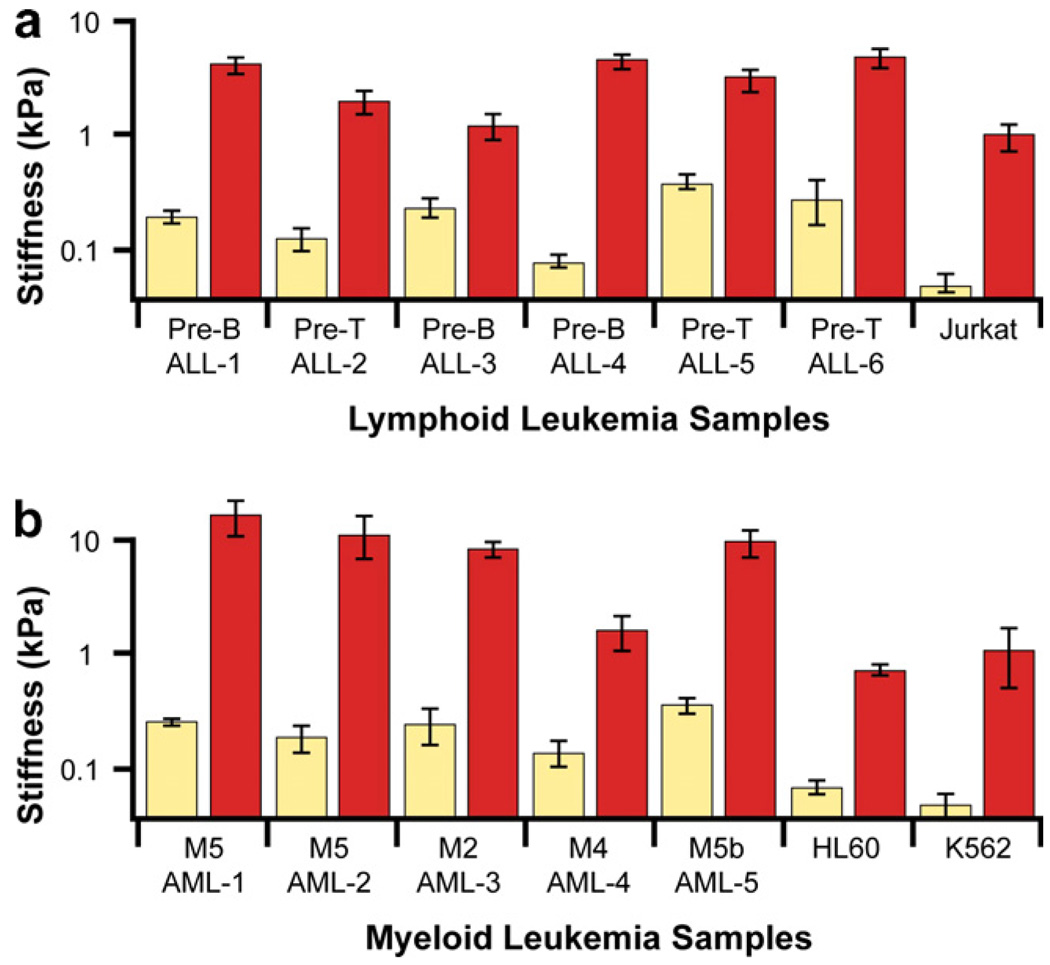
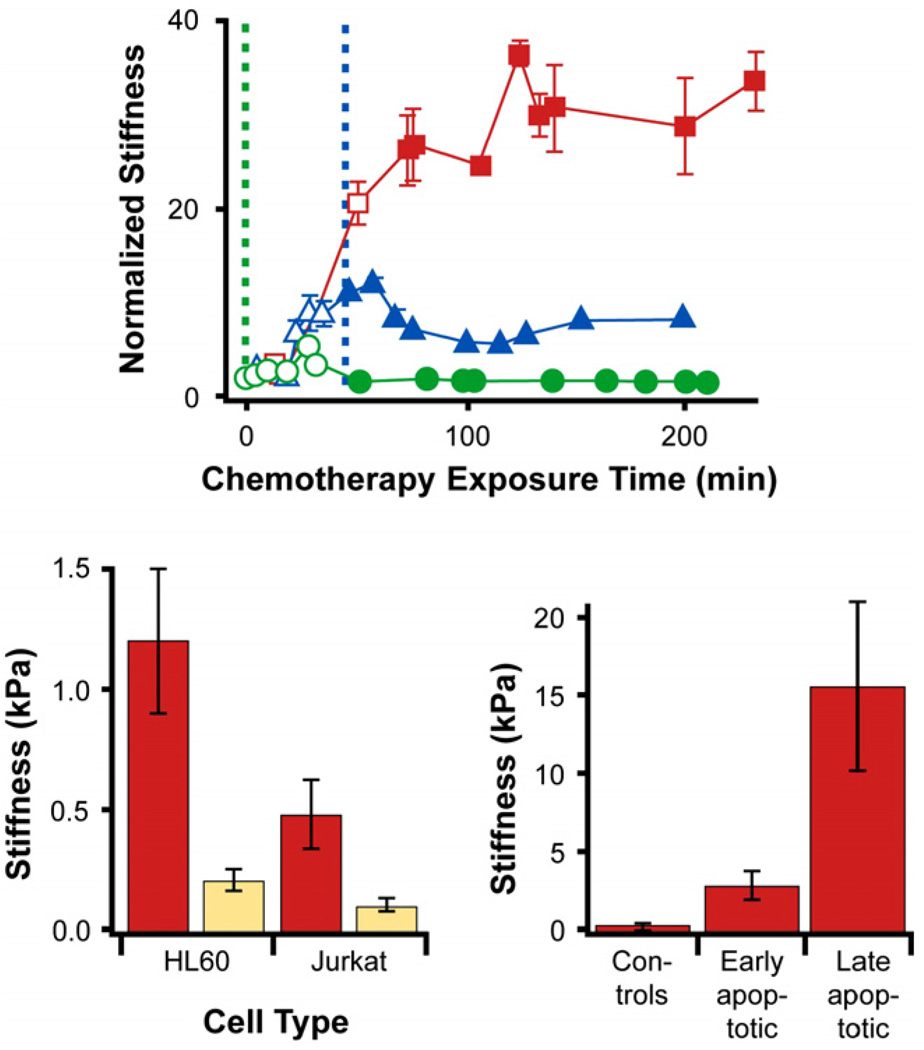
References
-
- Fung YC. Biomechanics: mechanical properties of living tissues. New York: Springer-Verlag; 1993.
-
- Fung YC. Biomechanics: motion, flow, stress, and growth. New York: Springer-Verlag; 1990.
-
- Fung YC. Biomechanics: circulation. 2nd ed. New York: Springer-Verlag; 1997.
-
- Fredberg JJ, Stamenovic D. On the imperfect elasticity of lung tissue. J Appl Physiol. 1989;67:2408–2419. - PubMed
-
- Humphrey JD. Cardiovascular solid mechanics: cells, tissues, organs. New York: Springer-Verlag; 2002.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
