

Sequential opening of mitochondrial ion channels as a function of glutathione redox thiol status
- PMID: 17540766
- PMCID: PMC2292488
- DOI: 10.1074/jbc.M702841200
Sequential opening of mitochondrial ion channels as a function of glutathione redox thiol status
Abstract
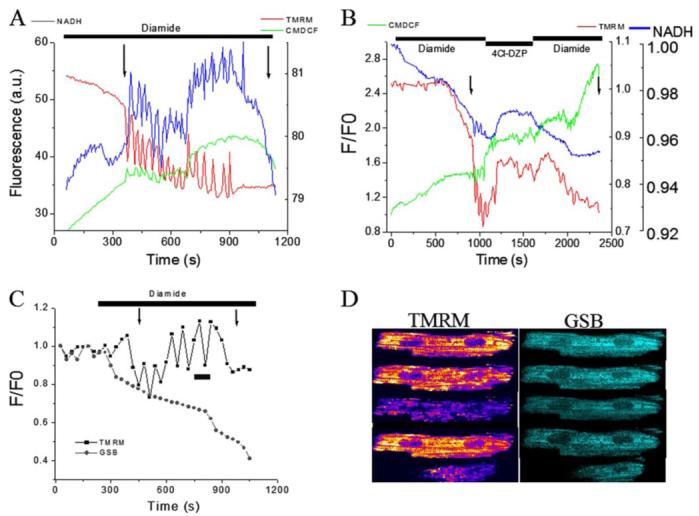
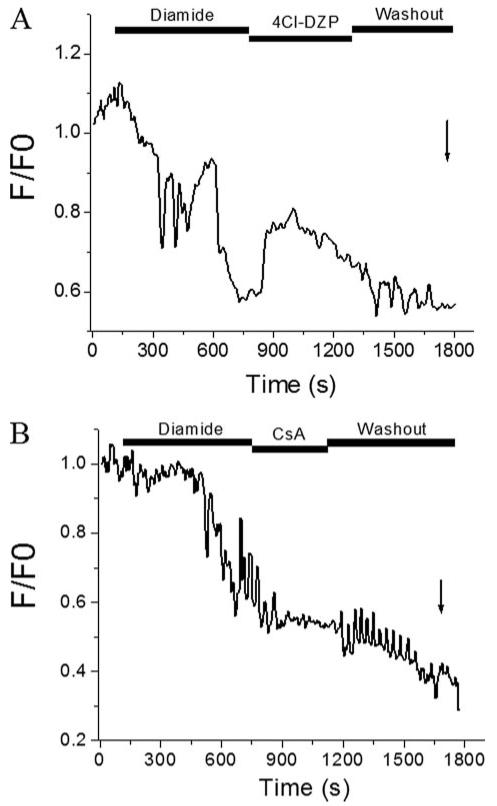
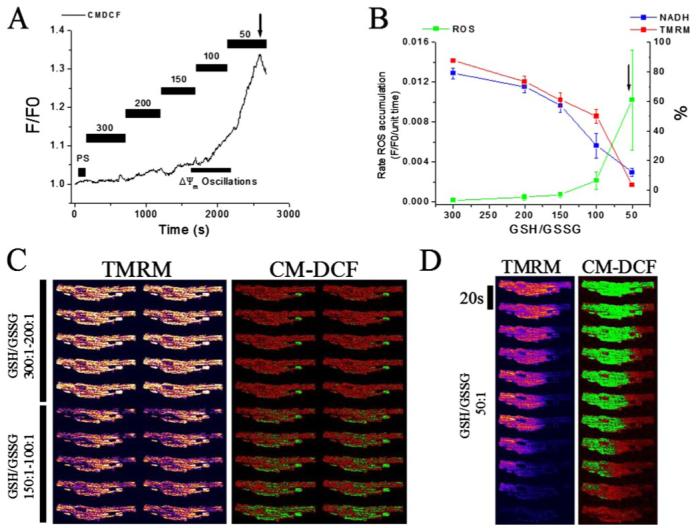
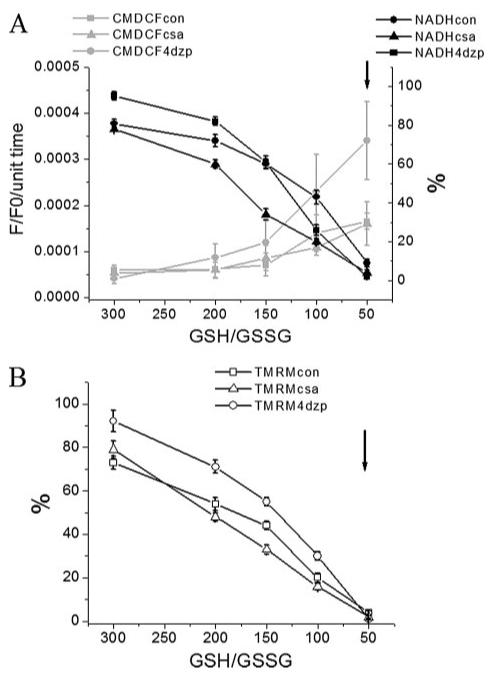
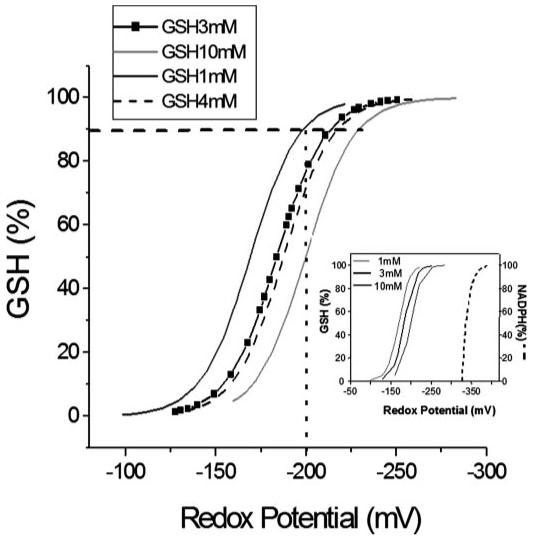
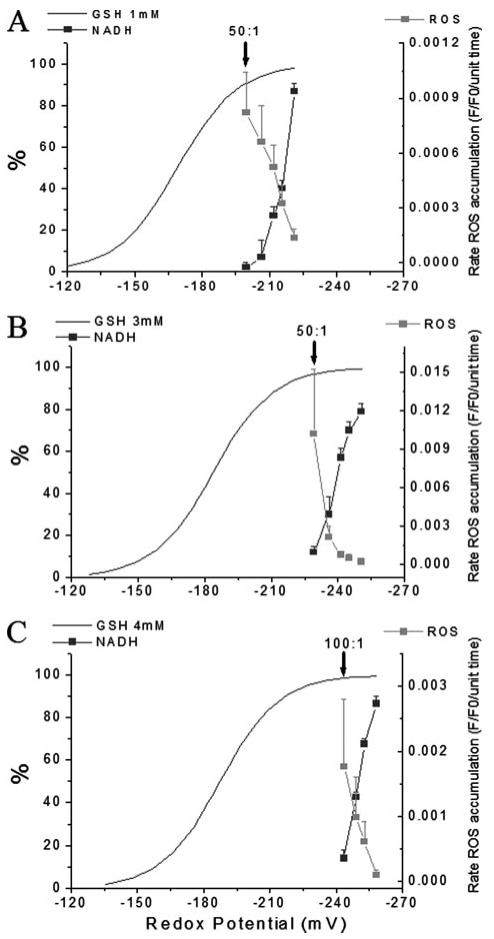
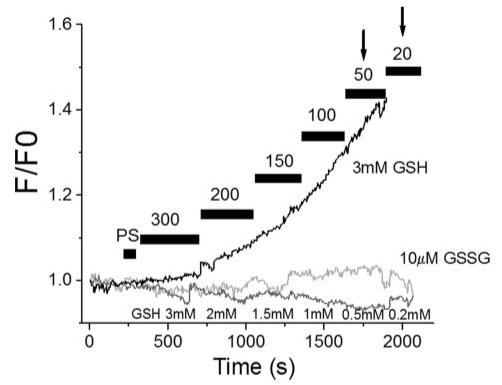
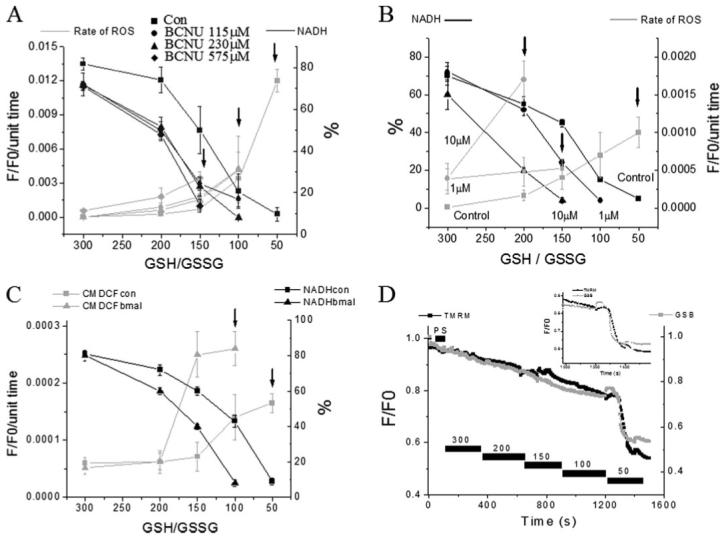
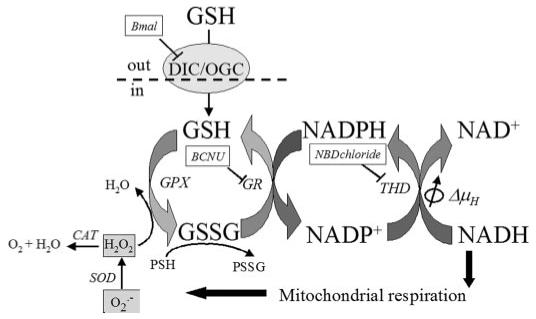
Mitochondrial membrane potential (DeltaPsi(m)) depolarization contributes to cell death and electrical and contractile dysfunction in the post-ischemic heart. An imbalance between mitochondrial reactive oxygen species production and scavenging was previously implicated in the activation of an inner membrane anion channel (IMAC), distinct from the permeability transition pore (PTP), as the first response to metabolic stress in cardiomyocytes. The glutathione redox couple, GSH/GSSG, oscillated in parallel with DeltaPsi(m) and the NADH/NAD(+) redox state. Here we show that depletion of reduced glutathione is an alternative trigger of synchronized mitochondrial oscillation in cardiomyocytes and that intermediate GSH/GSSG ratios cause reversible DeltaPsi(m) depolarization, although irreversible PTP activation is induced by extensive thiol oxidation. Mitochondrial dysfunction in response to diamide occurred in stages, progressing from oscillations in DeltaPsi(m) to sustained depolarization, in association with depletion of GSH. Mitochondrial oscillations were abrogated by 4'-chlorodiazepam, an IMAC inhibitor, whereas cyclosporin A was ineffective. In saponin-permeabilized cardiomyocytes, the thiol redox status was systematically clamped at GSH/GSSG ratios ranging from 300:1 to 20:1. At ratios of 150:1-100:1, DeltaPsi(m) depolarized reversibly, and a matrix-localized fluorescent marker was retained; however, decreasing the GSH/GSSG to 50:1 irreversibly depolarized DeltaPsi(m) and induced maximal rates of reactive oxygen species production, NAD(P)H oxidation, and loss of matrix constituents. Mitochondrial GSH sensitivity was altered by inhibiting either GSH uptake, the NADPH-dependent glutathione reductase, or the NADH/NADPH transhydrogenase, indicating that matrix GSH regeneration or replenishment was crucial. The results indicate that GSH/GSSG redox status governs the sequential opening of mitochondrial ion channels (IMAC before PTP) triggered by thiol oxidation in cardiomyocytes.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
