

POU-V factors antagonize maternal VegT activity and beta-Catenin signaling in Xenopus embryos
- PMID: 17541407
- PMCID: PMC1894774
- DOI: 10.1038/sj.emboj.7601736
POU-V factors antagonize maternal VegT activity and beta-Catenin signaling in Xenopus embryos
Abstract
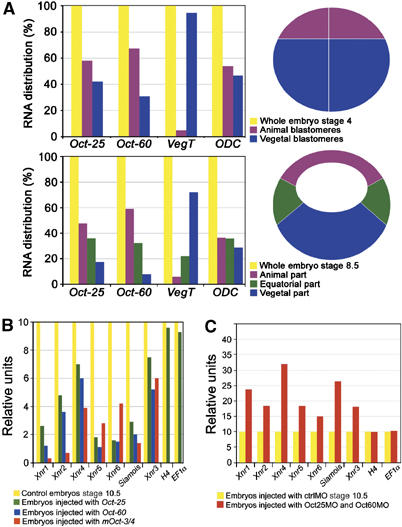
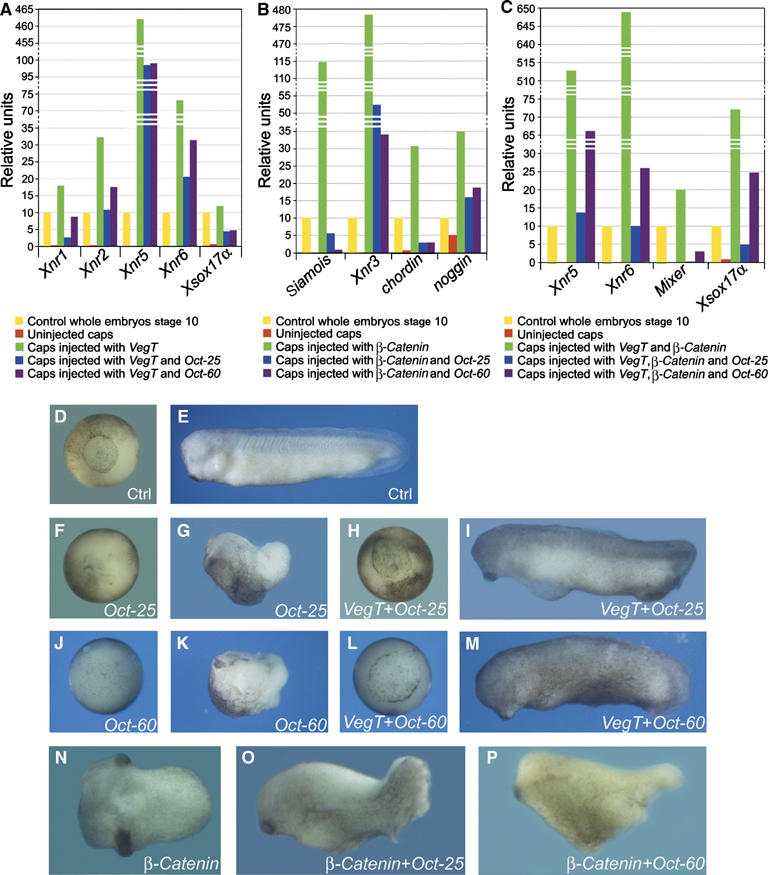
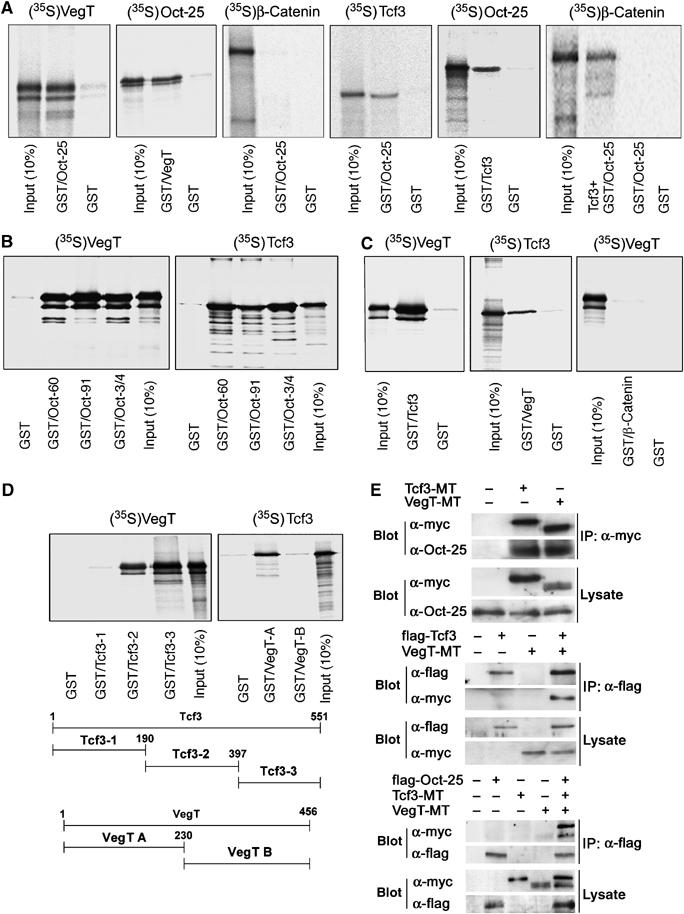
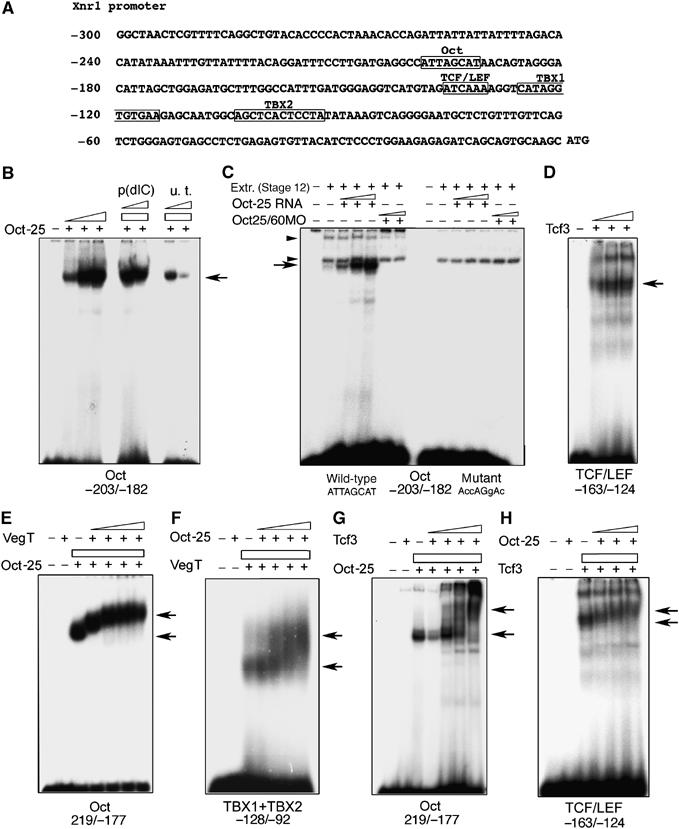
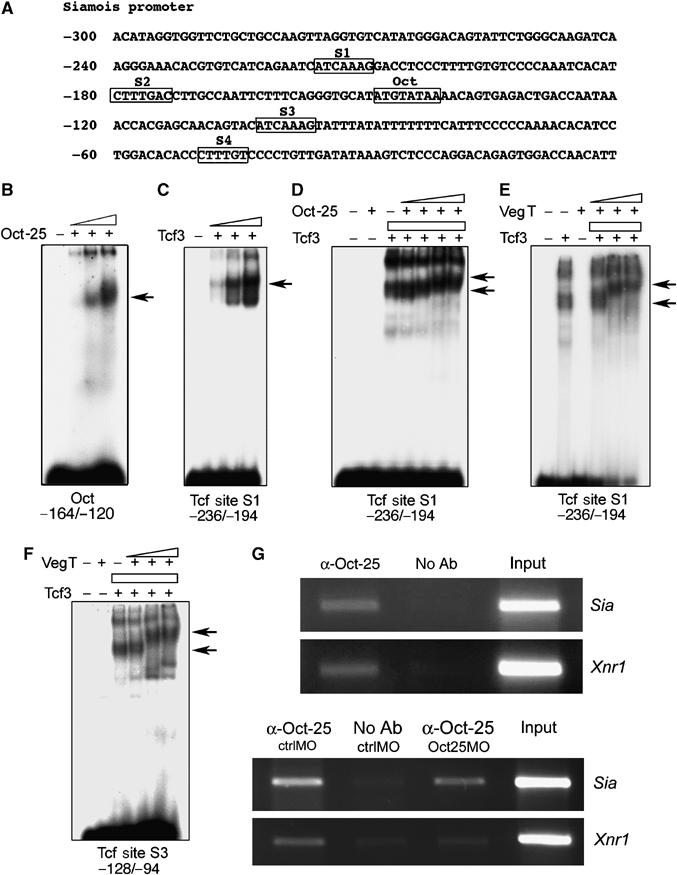
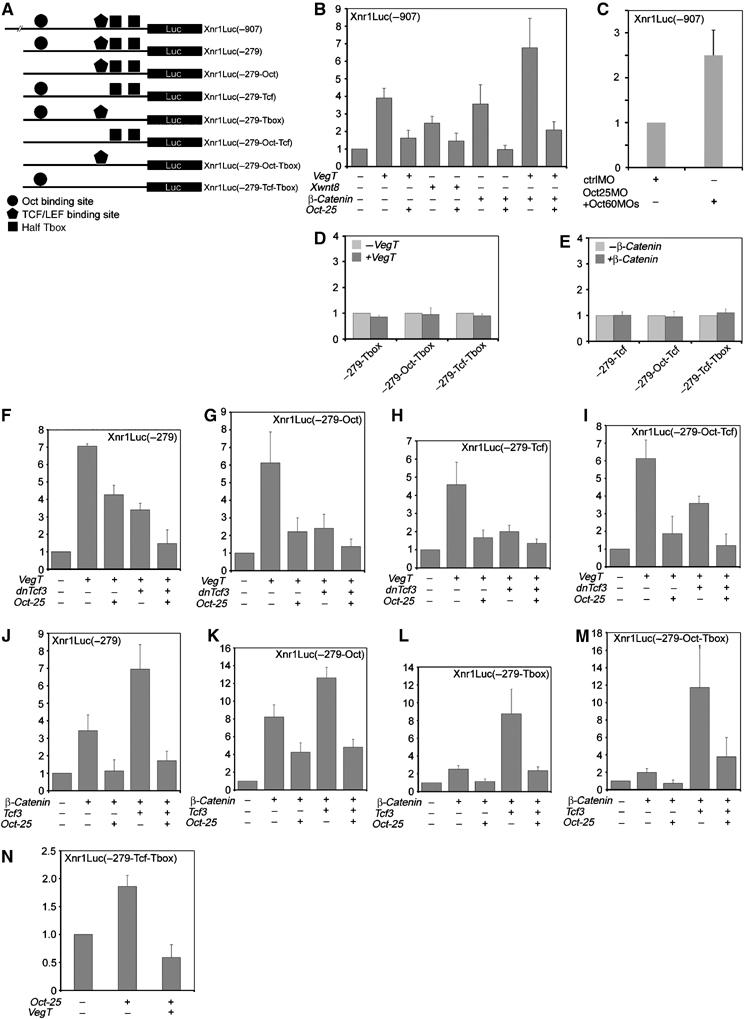
VegT and beta-Catenin are key players in the hierarchy of factors that are required for induction and patterning of mesendoderm in Xenopus embryogenesis. By descending the genetic cascades, cells lose their pluripotent status and are determined to differentiate into distinct tissues. Mammalian Oct-3/4, a POU factor of subclass V (POU-V), is required for the maintenance of pluripotency of embryonic stem cells. However, its molecular function within the early embryo is yet poorly understood. We here show that the two maternal Xenopus POU-V factors, Oct-60 and Oct-25, inhibit transcription of genes activated by VegT and beta-Catenin. Maternal POU-V factors and maternal VegT show an opposite distribution along the animal/vegetal axis. Oct-25, VegT and Tcf3 interact with each other and form repression complexes on promoters of VegT and beta-Catenin target genes. We suggest that POU-V factors antagonize primary inducers to allow germ layer specification in a temporally and spatially coordinated manner.
Figures

References
-
- Babaie Y, Herwig G, Greber B, Brink TC, Wruck W, Groth D, Lehrach H, Burdon T, Adjaye J (2007) Analysis of OCT4 dependent transcriptional networks regulating self renewal and pluripotency in human embryonic stem cells. Stem Cells 25: 500–510 - PubMed
-
- Behrens J, von Kries JP, Kühl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W (1996) Functional interaction of beta-Catenin with the transcription factor LEF-1. Nature 382: 638–642 - PubMed
-
- Boiani M, Schöler HR (2005) Regulatory networks in embryo derived pluripotent stem cells. Nat Rev Mol Cell Biol 6: 872–884 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
