

Essential role of lysyl oxidases in notochord development
- PMID: 17543297
- PMCID: PMC2467443
- DOI: 10.1016/j.ydbio.2007.04.029
Essential role of lysyl oxidases in notochord development
Abstract
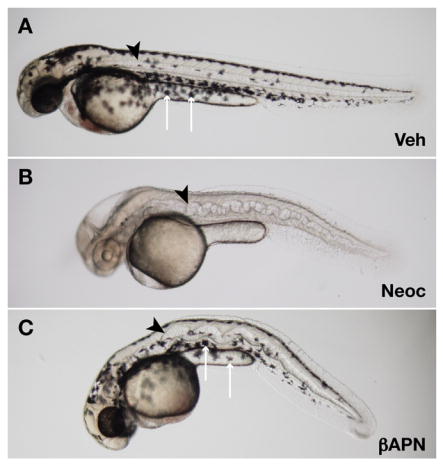
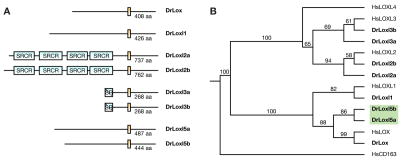
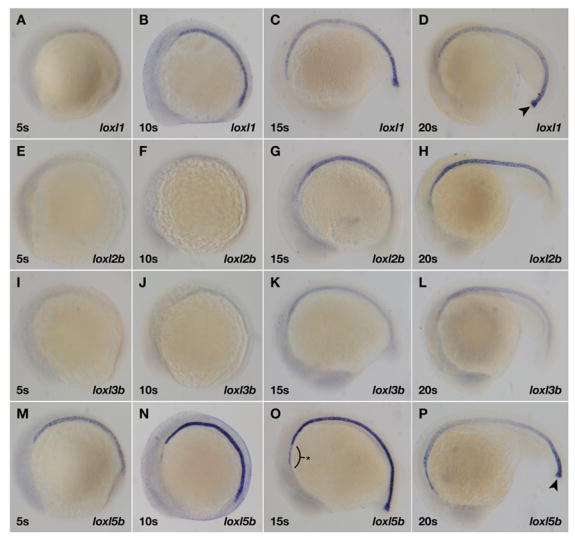
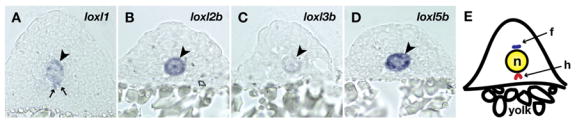
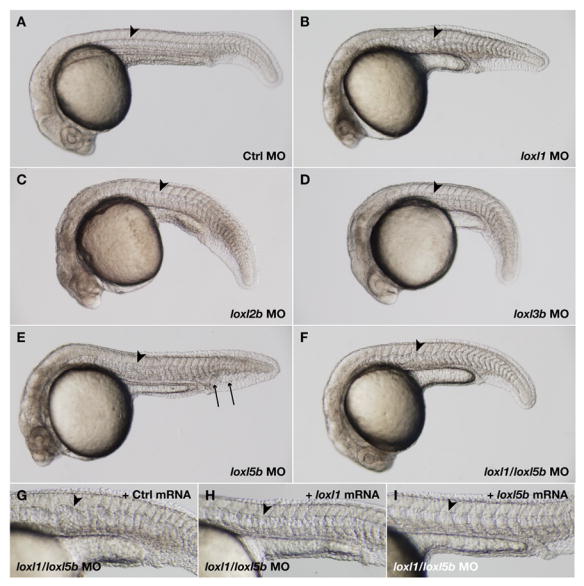
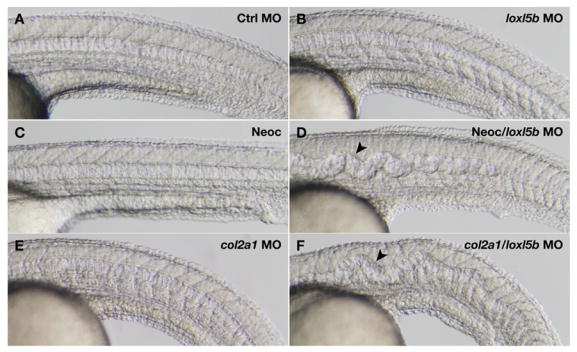
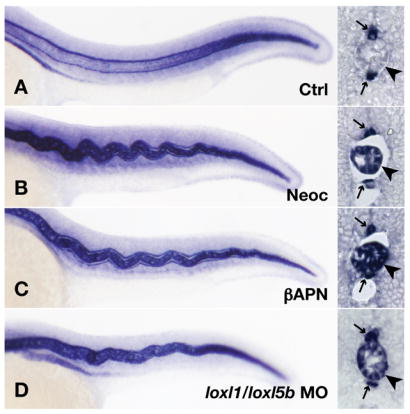
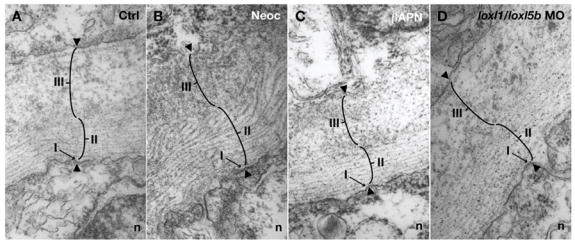
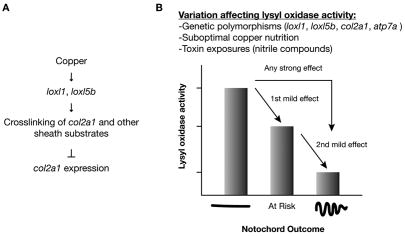
Recent studies reveal a critical role for copper in the development of the zebrafish notochord, suggesting that specific cuproenzymes are required for the structural integrity of the notochord sheath. We now demonstrate that beta-aminopropionitrile, a known inhibitor of the copper-dependent lysyl oxidases, causes notochord distortion in the zebrafish embryo identical to that seen in copper deficiency. Characterization of the zebrafish lysyl oxidase genes reveals eight unique sequences, several of which are expressed in the developing notochord. Specific gene knockdown demonstrates that loss of loxl1 results in notochord distortion, and that loxl1 and loxl5b have overlapping roles in notochord formation. Interestingly, while notochord abnormalities are not observed following partial knockdown of loxl1 or loxl5b alone, in each case this markedly sensitizes developing embryos to notochord distortion if copper availability is diminished. Likewise, partial knockdown of the lysyl oxidase substrate col2a1 results in notochord distortion when combined with reduced copper availability or partial knockdown of loxl1 or loxl5b. These data reveal a complex interplay of gene expression and nutrient availability critical to notochord development. They also provide insight into specific genetic and nutritional factors that may play a role in the pathogenesis of structural birth defects of the axial skeleton.
Figures
References
-
- Asuncion L, Fogelgren B, Fong KS, Fong SF, Kim Y, Csiszar K. A novel human lysyl oxidase-like gene (LOXL4) on chromosome 10q24 has an altered scavenger receptor cysteine rich domain. Matrix Biol. 2001;20:487–91. - PubMed
-
- Barrow MV, Simpson CF, Miller EJ. Lathyrism: a review. Q Rev Biol. 1974;49:101–28. - PubMed
-
- Bendtsen JD, Nielsen H, von Heijne G, Brunak S. Improved prediction of signal peptides: SignalP 3.0. J Mol Biol. 2004;340:783–95. - PubMed
-
- Borel A, Eichenberger D, Farjanel J, Kessler E, Gleyzal C, Hulmes DJ, Sommer P, Font B. Lysyl oxidase-like protein from bovine aorta. Isolation and maturation to an active form by bone morphogenetic protein-1. J Biol Chem. 2001;276:48944–9. - PubMed
-
- Cerda J, Grund C, Franke WW, Brand M. Molecular characterization of Calymmin, a novel notochord sheath-associated extracellular matrix protein in the zebrafish embryo. Dev Dyn. 2002;224:200–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
