

Functional redundancy of GSK-3alpha and GSK-3beta in Wnt/beta-catenin signaling shown by using an allelic series of embryonic stem cell lines
- PMID: 17543867
- PMCID: PMC4485918
- DOI: 10.1016/j.devcel.2007.04.001
Functional redundancy of GSK-3alpha and GSK-3beta in Wnt/beta-catenin signaling shown by using an allelic series of embryonic stem cell lines
Abstract
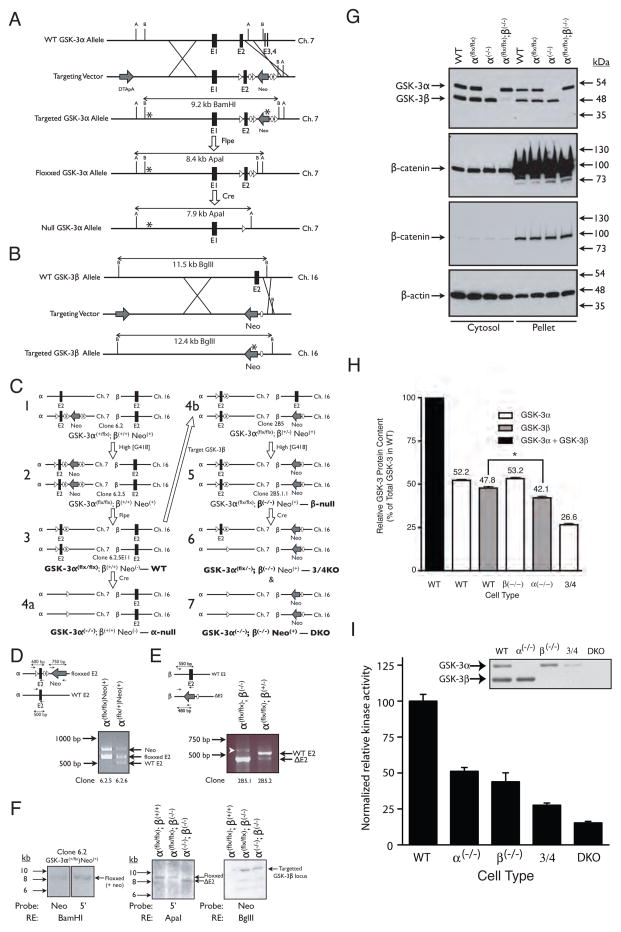
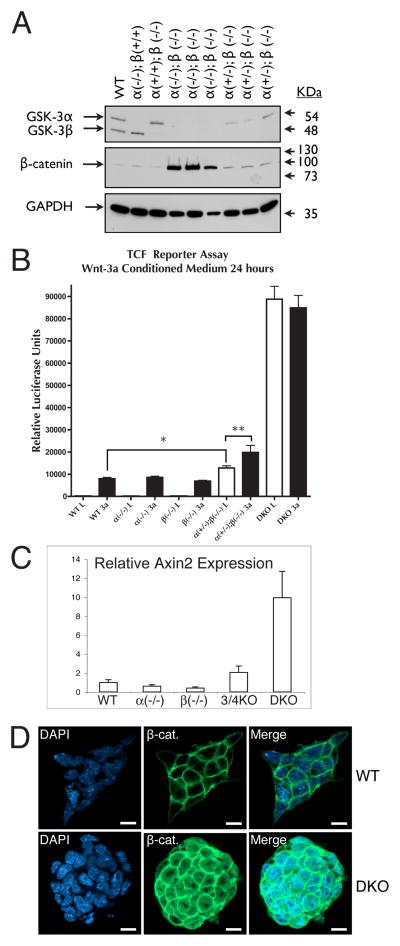
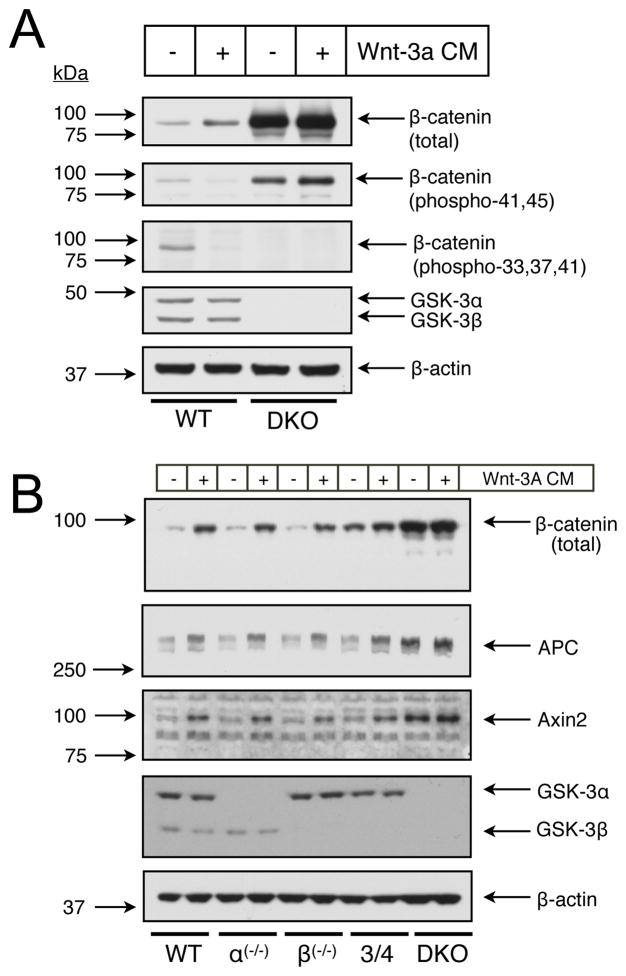
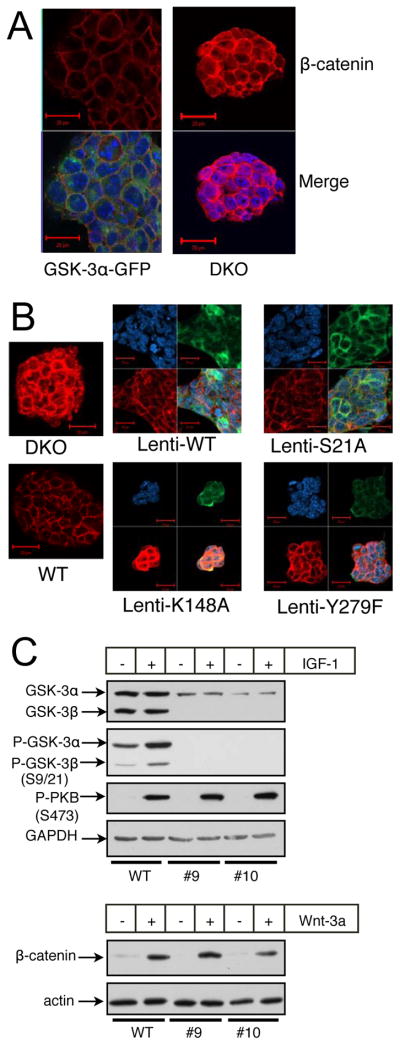
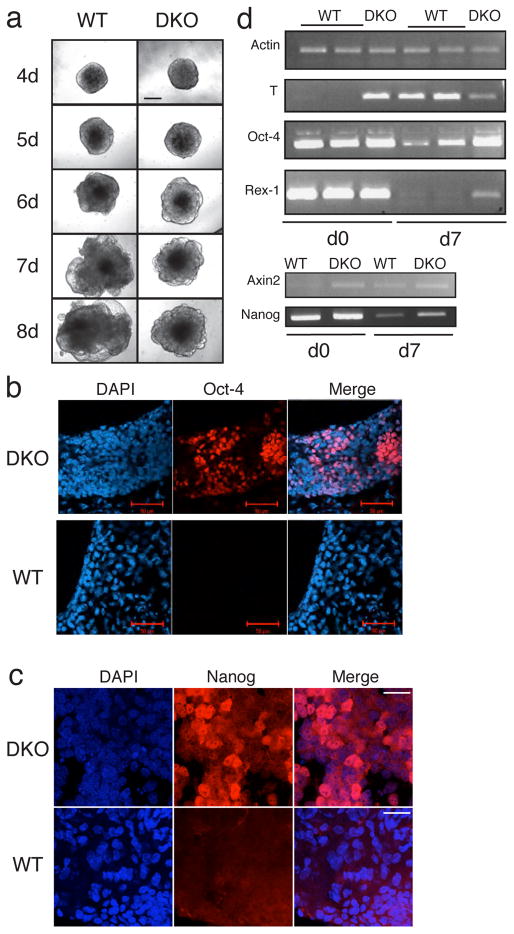
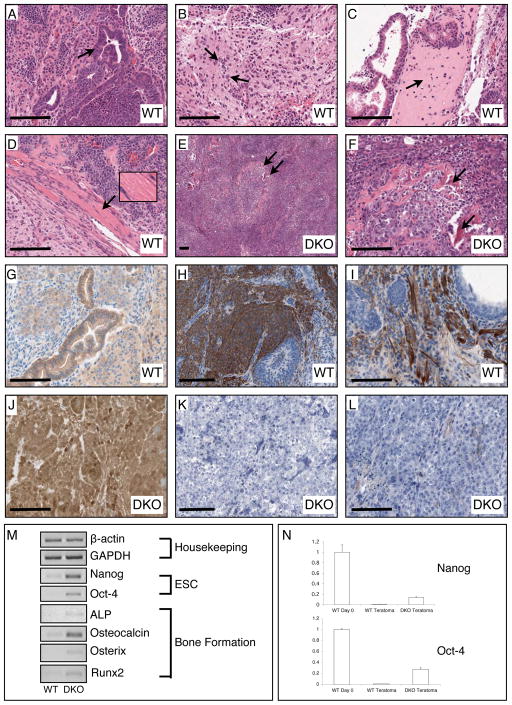
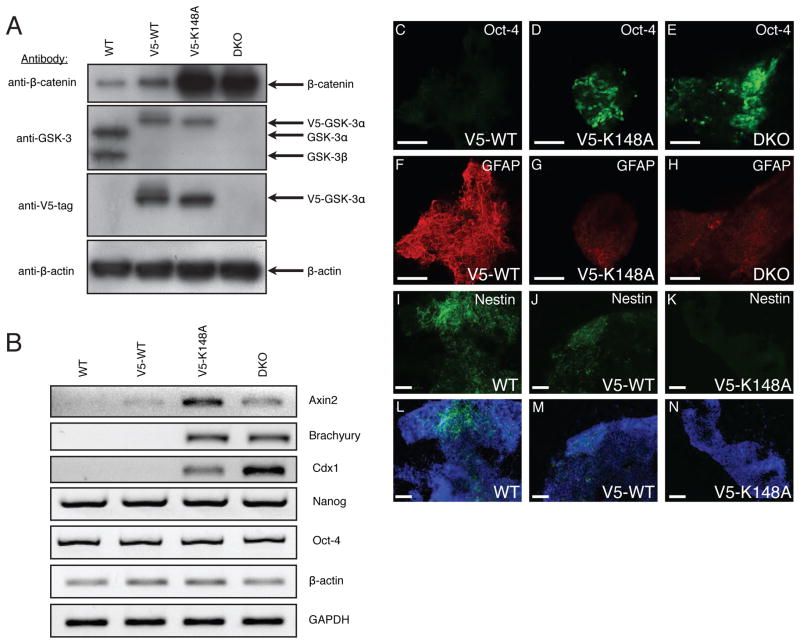
In mammalian cells, glycogen synthase kinase-3 (GSK-3) exists as two homologs, GSK-3alpha and GSK-3beta, encoded by independent genes, which share similar kinase domains but differ substantially in their termini. Here, we describe the generation of an allelic series of mouse embryonic stem cell (ESC) lines with 0-4 functional GSK-3 alleles and examine GSK-3-isoform function in Wnt/beta-catenin signaling. No compensatory upregulation in GSK-3 protein levels or activity was detected in cells lacking either GSK-3alpha or GSK-3beta, and Wnt/beta-catenin signaling was normal. Only in cells lacking three or all four of the alleles was a gene-dosage effect on beta-catenin/TCF-mediated transcription observed. Indeed, GSK-3alpha/beta double-knockout ESCs displayed hyperactivated Wnt/beta-catenin signaling and were severely compromised in their ability to differentiate, but could be rescued to normality by re-expression of functional GSK-3. The rheostatic regulation of GSK-3 highlights the importance of considering the contributions of both homologs when studying GSK-3 functions in mammalian systems.
Figures
References
-
- Arnold SJ, Stappert J, Bauer A, Kispert A, Herrmann BG, Kemler R. Brachyury is a target gene of the Wnt/beta-catenin signaling pathway. Mech Dev. 2000;91:249–258. - PubMed
-
- Bugeon L, Syed N, Dallman MJ. A fast and efficient method for transiently transfecting ES cells: application to the development of systems for conditional gene expression. Transgenic Res. 2000;9:229–232. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
