

Dopaminergic modulation of short-term synaptic plasticity at striatal inhibitory synapses
- PMID: 17545307
- PMCID: PMC1885397
- DOI: 10.1073/pnas.0703813104
Dopaminergic modulation of short-term synaptic plasticity at striatal inhibitory synapses
Abstract
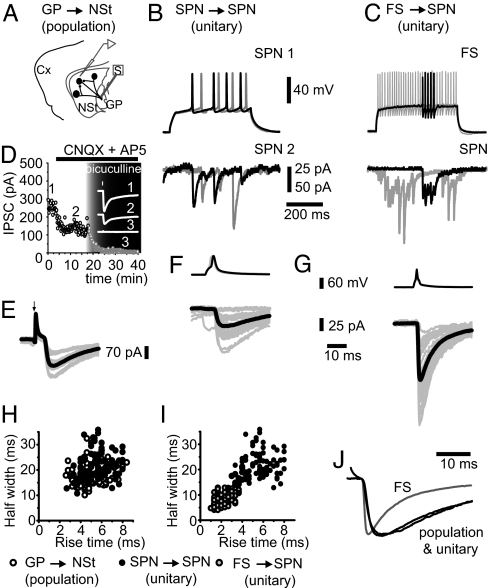
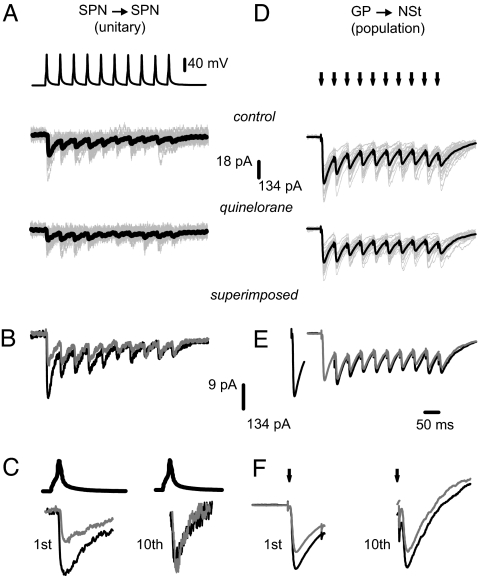
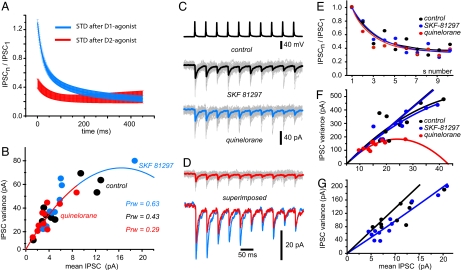
Circuit properties, such as the selection of motor synergies, have been posited as relevant tasks for the recurrent inhibitory synapses between spiny projection neurons of the neostriatum, a nucleus of the basal ganglia participating in procedural learning and voluntary motor control. Here we show how the dopaminergic system regulates short-term plasticity (STP) in these synapses. STP is thought to endow neuronal circuits with computational powers such as gain control, filtering, and the emergence of transitory net states. But little is known about STP regulation. Employing unitary and population synaptic recordings, we observed that activation of dopamine receptors can modulate STP between spiny neurons. A D(1)-class agonist enhances, whereas a D(2)-class agonist decreases, short-term depression most probably by synaptic redistribution. Presynaptic receptors appear to be responsible for this modulation. In contrast, STP between fast-spiking interneurons and spiny projection neurons is largely unregulated despite expressing presynaptic receptors. Thus, the present experiments provide an explanation for dopamine actions at the circuit level: the control of STP between lateral connections of output neurons and the reorganization of the balance between different forms of inhibitory transmission. Theoretically, D(1) receptors would promote a sensitive, responsive state for temporal precision (dynamic component), whereas D(2) receptors would sense background activity (static component).
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
