

NCgl2620 encodes a class II polyphosphate kinase in Corynebacterium glutamicum
- PMID: 17545325
- PMCID: PMC1951008
- DOI: 10.1128/AEM.00600-07
NCgl2620 encodes a class II polyphosphate kinase in Corynebacterium glutamicum
Abstract
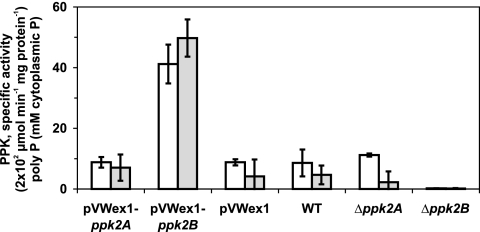
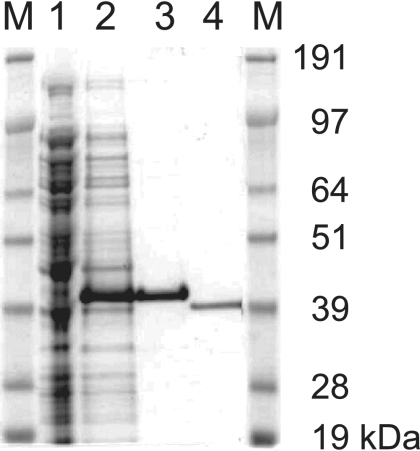
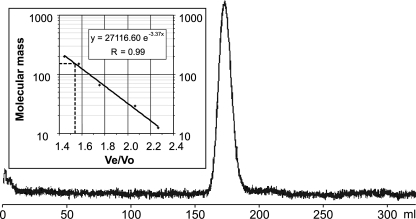
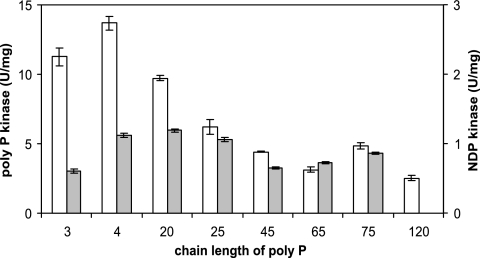
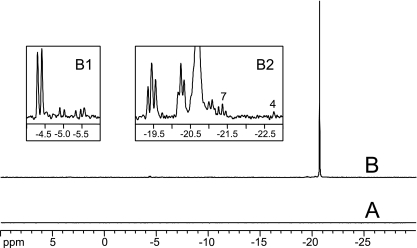
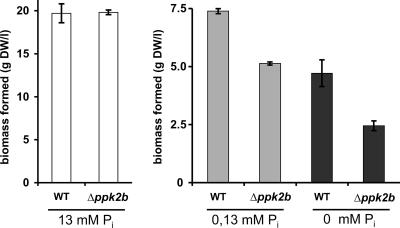
Corynebacterium glutamicum is able to accumulate up to 600 mM cytosolic phosphorus in the form of polyphosphate (poly P). Granular poly P (volutin) can make up to 37% of the internal cell volume. This bacterium lacks the classic enzyme of poly P synthesis, class I polyphosphate kinase (PPK1), but it possesses two genes, ppk2A (corresponds to NCgl0880) and ppk2B (corresponds to NCgl2620), for putative class II (PPK2) PPKs. Deletion of ppk2B decreased PPK activity and cellular poly P content, while overexpression of ppk2B increased both PPK activity and cellular poly P content. Neither deletion nor overexpression of ppk2A changed specific activity of PPK or cellular poly P content significantly. Purified PPK2B of C. glutamicum is active as a homotetramer and formed poly P with an average chain length of about 125, as determined with (31)P nuclear magnetic resonance. The catalytic efficiency of C. glutamicum PPK2B was higher in the poly P-forming direction than for nucleoside triphosphate formation from poly P. The ppk2B deletion mutant, which accumulated very little poly P and grew as C. glutamicum wild type under phosphate-sufficient conditions, showed a growth defect under phosphate-limiting conditions.
Figures
References
-
- Abe, S. G.-G., K. I. Takayama, and S. Kinoshita. 1967. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 13:279-285.
-
- Ahn, K., and A. Kornberg. 1990. Polyphosphate kinase from Escherichia coli. Purification and demonstration of a phosphoenzyme intermediate. J. Biol. Chem. 265:11734-11739. - PubMed
-
- Akiyama, M., E. Crooke, and A. Kornberg. 1992. The polyphosphate kinase gene of Escherichia coli. Isolation and sequence of the ppk gene and membrane location of the protein. J. Biol. Chem. 267:22556-22561. - PubMed
-
- Ayraud, S., B. Janvier, A. Labigne, C. Ecobichon, C. Burucoa, and J. L. Fauchere. 2005. Polyphosphate kinase: a new colonization factor of Helicobacter pylori. FEMS Microbiol. Lett. 243:45-50. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
