

Alpha4beta1 integrin and erythropoietin mediate temporally distinct steps in erythropoiesis: integrins in red cell development
- PMID: 17548514
- PMCID: PMC2064286
- DOI: 10.1083/jcb.200702080
Alpha4beta1 integrin and erythropoietin mediate temporally distinct steps in erythropoiesis: integrins in red cell development
Erratum in
- J Cell Biol. 2008 Apr 21;181(2):395
Abstract
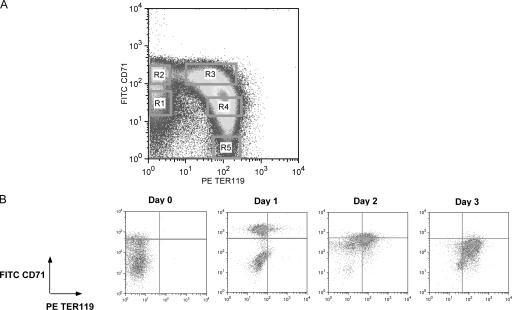
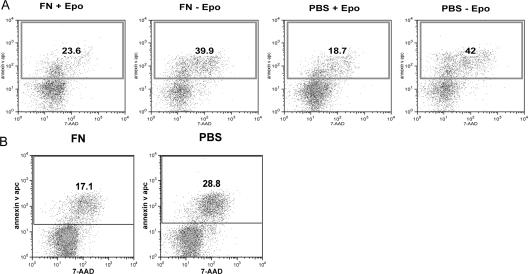
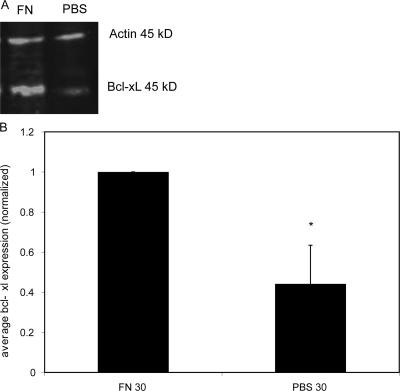
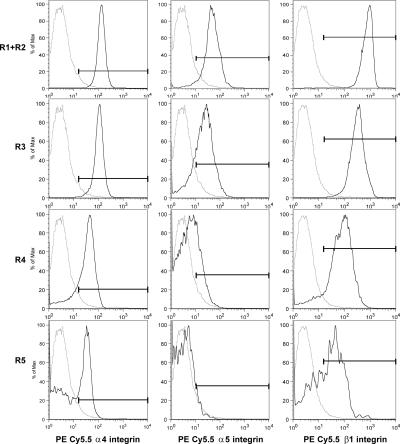
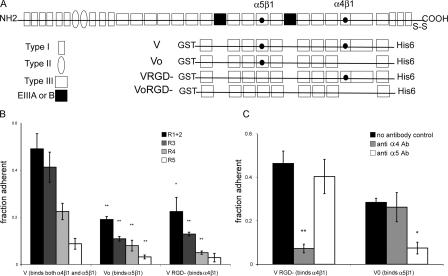
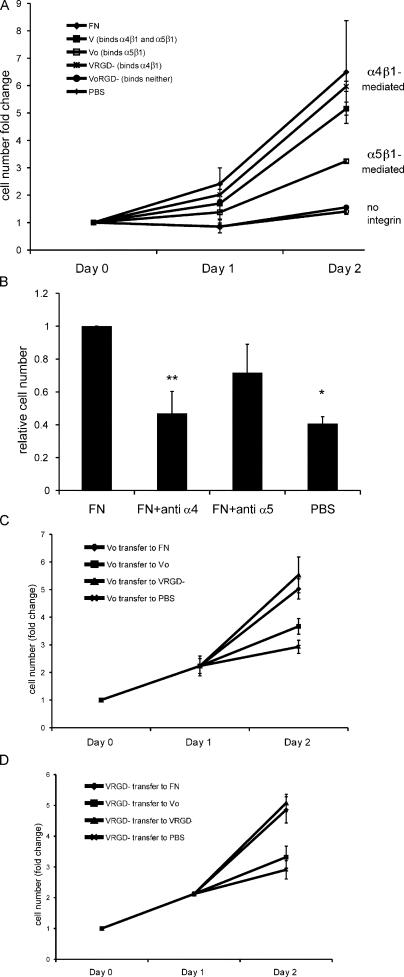
Erythropoietin (Epo) is essential for the terminal proliferation and differentiation of erythroid progenitor cells. Fibronectin is an important part of the erythroid niche, but its precise role in erythropoiesis is unknown. By culturing fetal liver erythroid progenitors, we show that fibronectin and Epo regulate erythroid proliferation in temporally distinct steps: an early Epo-dependent phase is followed by a fibronectin-dependent phase. In each phase, Epo and fibronectin promote expansion by preventing apoptosis partly through bcl-xL. We show that alpha(4), alpha(5), and beta(1) are the principal integrins expressed on erythroid progenitors; their down-regulation during erythropoiesis parallels the loss of cell adhesion to fibronectin. Culturing erythroid progenitors on recombinant fibronectin fragments revealed that only substrates that engage alpha(4)beta(1)-integrin support normal proliferation. Collectively, these data suggest a two-phase model for growth factor and extracellular matrix regulation of erythropoiesis, with an early Epo-dependent, integrin-independent phase followed by an Epo-independent, alpha(4)beta(1)-integrin-dependent phase.
Figures




References
-
- Arroyo, A.G., J.T. Yang, H. Rayburn, and R.O. Hynes. 1999. Alpha4 integrins regulate the proliferation/differentiation balance of multilineage hematopoietic progenitors in vivo. Immunity. 11:555–566. - PubMed
-
- Avraham, H., S.Y. Park, K. Schinkmann, and S. Avraham. 2000. RAFTK/Pyk2-mediated cellular signalling. Cell. Signal. 12:123–133. - PubMed
-
- Campos, L.S. 2005. Beta1 integrins and neural stem cells: making sense of the extracellular environment. Bioessays. 27:698–707. - PubMed
-
- Fawcett, D.W., and R.P. Jensh. 1997. Bloom & Fawcett: Concise Histology. Chapman and Hall, New York. 84–93 pp.
-
- Gregory, C.J., and A.C. Eaves. 1977. Human marrow cells capable of erythropoietic differentiation in vitro: definition of three erythroid colony responses. Blood. 49:855–864. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
