

Contact-dependent inhibition of EGFR signaling by Nf2/Merlin
- PMID: 17548515
- PMCID: PMC2064288
- DOI: 10.1083/jcb.200703010
Contact-dependent inhibition of EGFR signaling by Nf2/Merlin
Abstract
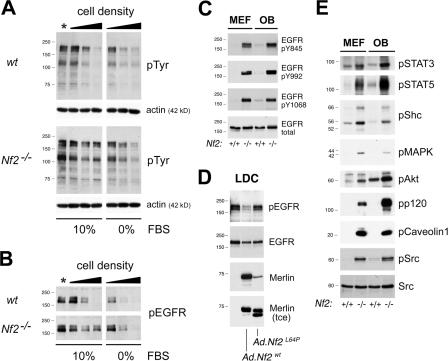
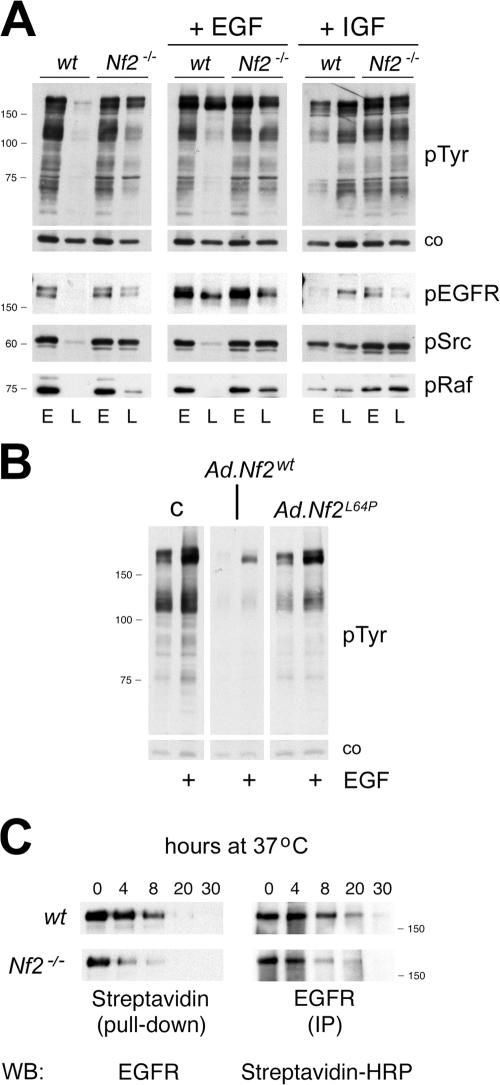
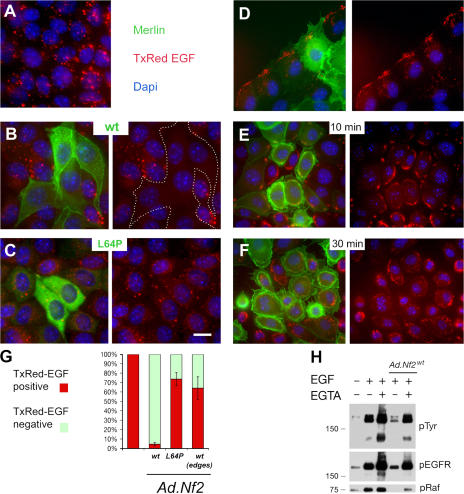
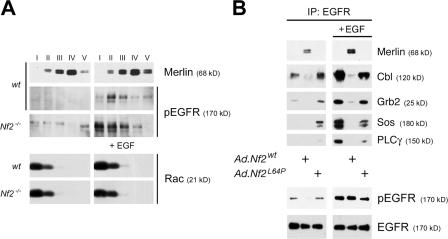
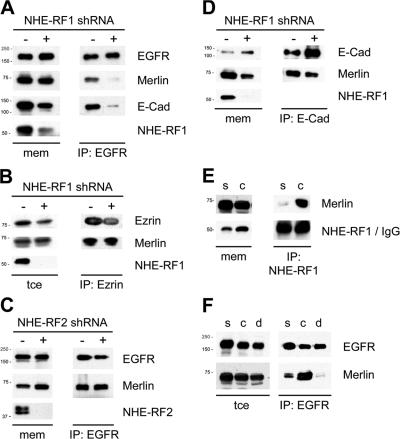
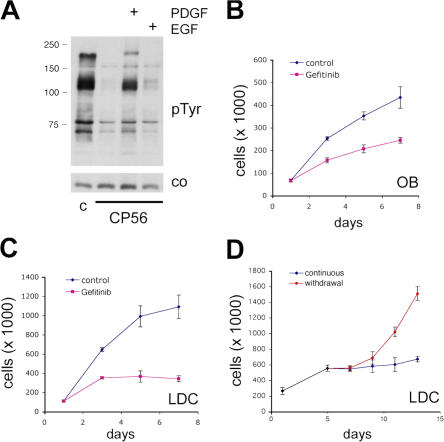
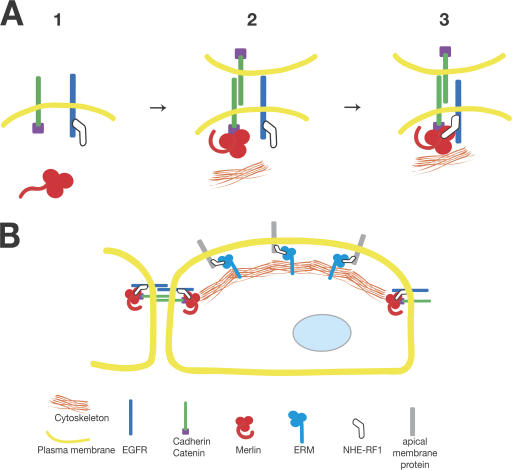
The neurofibromatosis type 2 (NF2) tumor suppressor, Merlin, is a membrane/cytoskeleton-associated protein that mediates contact-dependent inhibition of proliferation. Here we show that upon cell-cell contact Merlin coordinates the processes of adherens junction stabilization and negative regulation of epidermal growth factor receptor (EGFR) signaling by restraining the EGFR into a membrane compartment from which it can neither signal nor be internalized. In confluent Nf2(-/-) cells, EGFR activation persists, driving continued proliferation that is halted by specific EGFR inhibitors. These studies define a new mechanism of tumor suppression, provide mechanistic insight into the poorly understood phenomenon of contact-dependent inhibition of proliferation, and suggest a therapeutic strategy for NF2-mutant tumors.
Figures
References
-
- Bailey, S.N., S.M. Ali, A.E. Carpenter, C.O. Higgins, and D.M. Sabatini. 2006. Microarrays of lentiviruses for gene function screens in immortalized and primary cells. Nat. Methods. 3:117–122. - PubMed
-
- Baser, M.E., J.M. Friedman, A.J. Wallace, R.T. Ramsden, H. Joe, and D.G. Evans. 2002. Evaluation of clinical diagnostic criteria for neurofibromatosis 2. Neurology. 59:1759–1765. - PubMed
-
- Bennett, F.C., and K.F. Harvey. 2006. Fat cadherin modulates organ size in Drosophila via the Salvador/Warts/Hippo signaling pathway. Curr. Biol. 16:2101–2110. - PubMed
-
- Betson, M., E. Lozano, J. Zhang, and V.M. Braga. 2002. Rac activation upon cell-cell contact formation is dependent on signaling from the epidermal growth factor receptor. J. Biol. Chem. 277:36962–36969. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
