

Altering the distribution of Foxp3(+) regulatory T cells results in tissue-specific inflammatory disease
- PMID: 17548521
- PMCID: PMC2118615
- DOI: 10.1084/jem.20070081
Altering the distribution of Foxp3(+) regulatory T cells results in tissue-specific inflammatory disease
Abstract
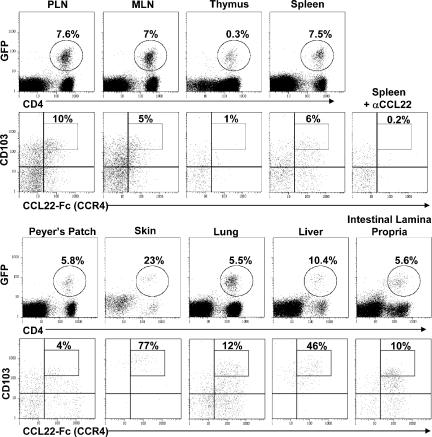
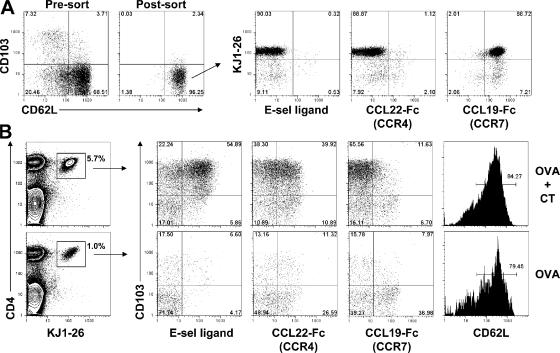
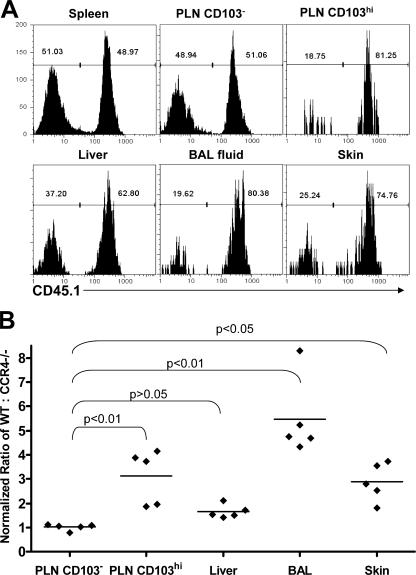
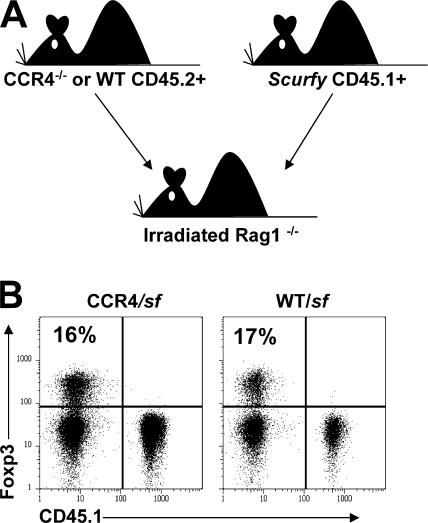
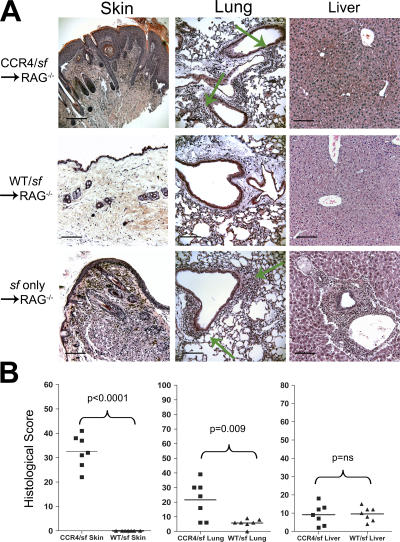
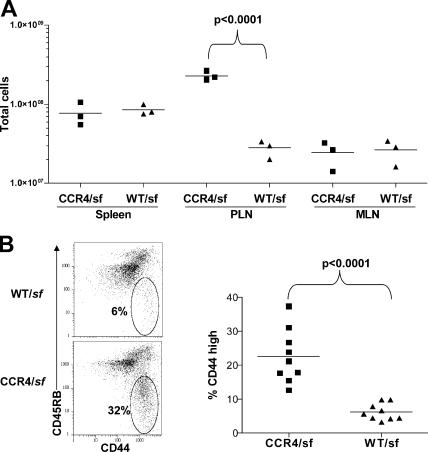
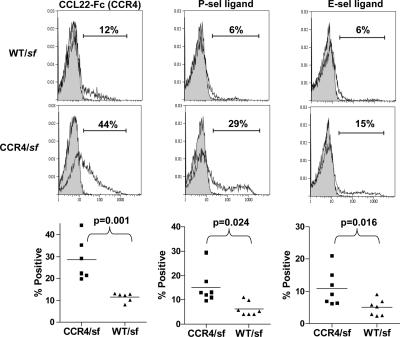
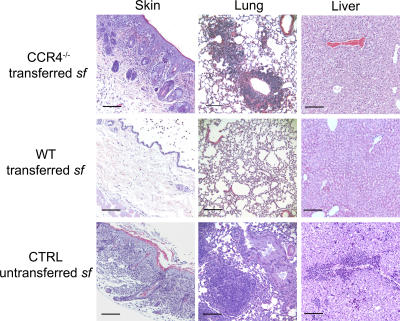
CD4(+)Foxp3(+) regulatory T cells (T reg) are essential for maintaining self-tolerance, but their functional mechanisms and sites of action in vivo are poorly defined. We examined the homing receptor expression and tissue distribution of T reg cells in the steady state and determined whether altering their distribution by removal of a single chemokine receptor impairs their ability to maintain tissue-specific peripheral tolerance. We found that T reg cells are distributed throughout all nonlymphoid tissues tested, and are particularly prevalent in the skin, where they express a unique CCR4(+)CD103(hi) phenotype. T reg cell expression of CCR4 and CD103 is induced by antigen-driven activation within subcutaneous lymph nodes, and accumulation of T reg cells in the skin and lung airways is impaired in the absence of CCR4 expression. Mice with a complete loss of CCR4 in the T reg cell compartment develop lymphocytic infiltration and severe inflammatory disease in the skin and lungs, accompanied by peripheral lymphadenopathy and increased differentiation of skin-tropic CD4(+)Foxp3(+) T cells. Thus, selectively altering T reg cell distribution in vivo leads to the development of tissue-specific inflammatory disease.
Figures
References
-
- Sakaguchi, S. 2004. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 22:531–562. - PubMed
-
- Szanya, V., J. Ermann, C. Taylor, C. Holness, and C.G. Fathman. 2002. The subpopulation of CD4+CD25+ splenocytes that delays adoptive transfer of diabetes expresses L-selectin and high levels of CCR7. J. Immunol. 169:2461–2465. - PubMed
-
- Salomon, B., D.J. Lenschow, L. Rhee, N. Ashourian, B. Singh, A. Sharpe, and J.A. Bluestone. 2000. B7/CD28 costimulation is essential for the homeostasis of the CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes. Immunity. 12:431–440. - PubMed
-
- Kohm, A.P., P.A. Carpentier, H.A. Anger, and S.D. Miller. 2002. Cutting edge: CD4+CD25+ regulatory T cells suppress antigen-specific autoreactive immune responses and central nervous system inflammation during active experimental autoimmune encephalomyelitis. J. Immunol. 169:4712–4716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
