

Protein kinase Cdelta regulates antigen receptor-induced lytic granule polarization in mouse CD8+ CTL
- PMID: 17548619
- PMCID: PMC3712856
- DOI: 10.4049/jimmunol.178.12.7814
Protein kinase Cdelta regulates antigen receptor-induced lytic granule polarization in mouse CD8+ CTL
Abstract
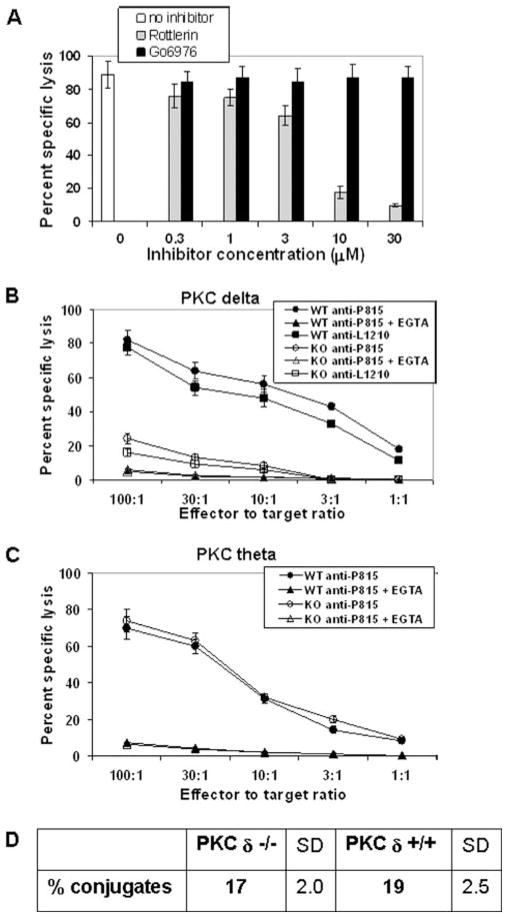
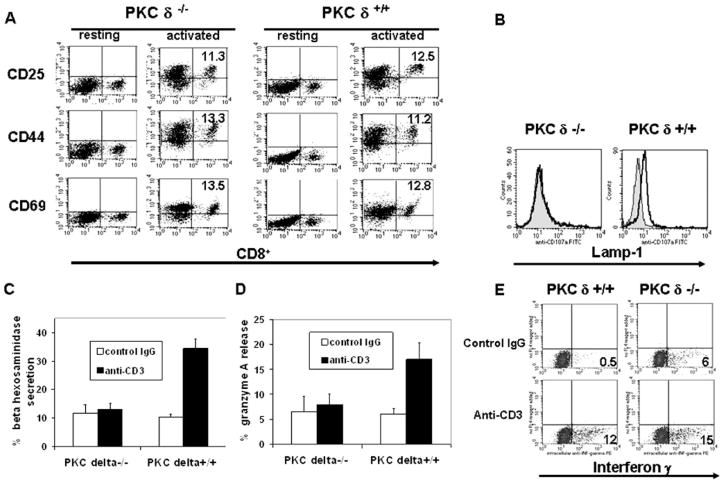
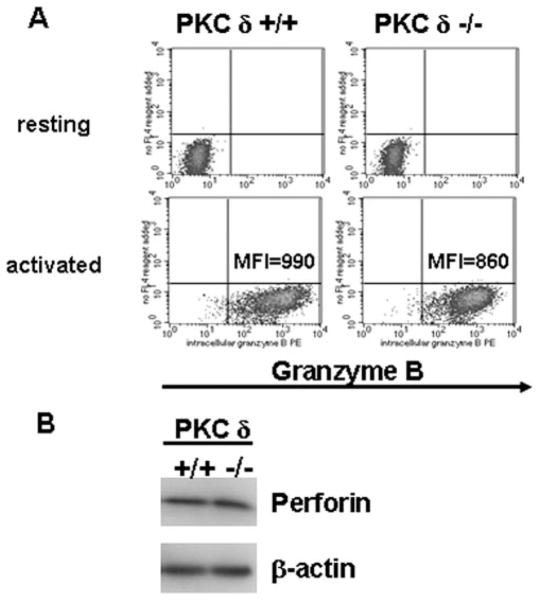
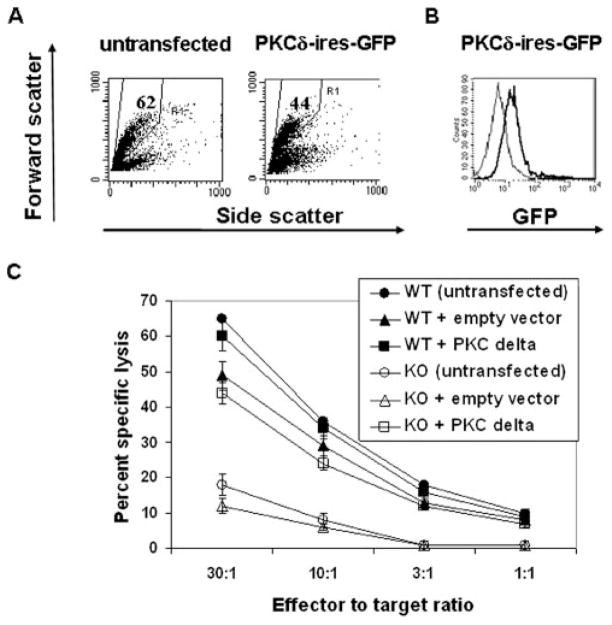
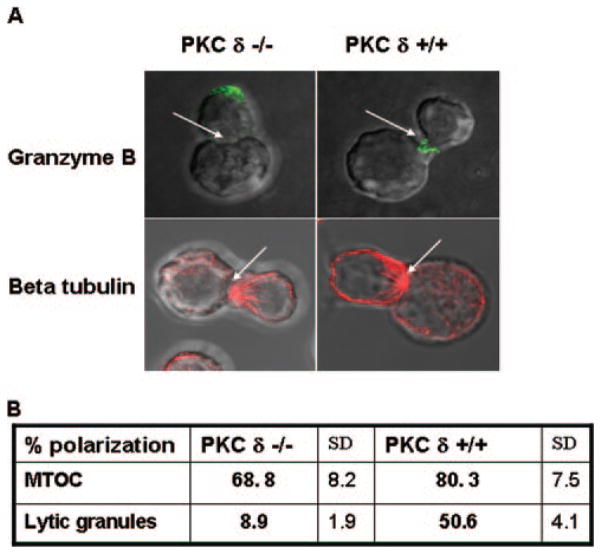
Lytic granule exocytosis is the major pathway used by CD8+ CTL to kill virally infected and tumor cells. Despite the obvious importance of this pathway in adaptive T cell immunity, the molecular identity of enzymes involved in the regulation of this process is poorly characterized. One signal known to be critical for the regulation of granule exocytosis-mediated cytotoxicity in CD8+ T cells is Ag receptor-induced activation of protein kinase C (PKC). However, it is not known which step of the process is regulated by PKC. In addition, it has not been determined to date which of the PKC family members is required for the regulation of lytic granule exocytosis. By combination of pharmacological inhibitors and use of mice with targeted gene deletions, we show that PKCdelta is required for granule exocytosis-mediated lytic function in mouse CD8+ T cells. Our studies demonstrate that PKCdelta is required for lytic granule exocytosis, but is dispensable for activation, cytokine production, and expression of cytolytic molecules in response to TCR stimulation. Importantly, defective lytic function in PKCdelta-deficient cytotoxic lymphocytes is reversed by ectopic expression of PKCdelta. Finally, we show that PKCdelta is not involved in target cell-induced reorientation of the microtubule-organizing center, but is required for the subsequent exocytosis step, i.e., lytic granule polarization. Thus, our studies identify PKCdelta as a novel and selective regulator of Ag receptor-induced lytic granule polarization in mouse CD8+ T cells.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
References
-
- Russell JH, Ley TJ. Lymphocyte-mediated cytotoxicity. Annu Rev Immunol. 2002;20:323–370. - PubMed
-
- Kagi D, Ledermann B, Burki K, Zinkernagel RM, Hengartner H. Molecular mechanisms of lymphocyte-mediated cytotoxicity and their role in immunological protection and pathogenesis in vivo. Annu Rev Immunol. 1996;14:207–302. - PubMed
-
- Berry M, Bleackley R. Cytotoxic lymphocytes: all roads lead to death. Nat Rev Immunol. 2002;2:401–409. - PubMed
-
- Blott EJ, Griffiths GM. Secretory lysosomes. Nat Rev Mol Cell Biol. 2002;3:122–131. - PubMed
-
- Lancki DW, Weiss A, Fitch FW. Requirements for triggering of lysis by cytolytic T lymphocyte clones. J Immunol. 1987;138:3646–3653. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
