

A homodimeric complex of HLA-G on normal trophoblast cells modulates antigen-presenting cells via LILRB1
- PMID: 17549736
- PMCID: PMC2699429
- DOI: 10.1002/eji.200737089
A homodimeric complex of HLA-G on normal trophoblast cells modulates antigen-presenting cells via LILRB1
Abstract
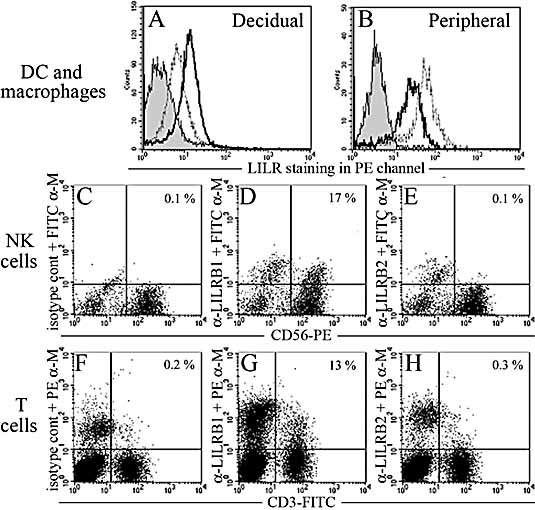
In healthy individuals, the non-classical MHC molecule HLA-G is only expressed on fetal trophoblast cells that invade the decidua during placentation. We show that a significant proportion of HLA-G at the surface of normal human trophoblast cells is present as a disulphide-linked homodimer of the conventional beta(2)m-associated HLA-I complex. HLA-G is a ligand for leukocyte immunoglobulin-like receptors (LILR), which bind much more efficiently to dimeric HLA-G than to conventional HLA-I molecules. We find that a LILRB1-Fc fusion protein preferentially binds the dimeric form of HLA-G on trophoblast cells. We detect LILRB1 expression on decidual myelomonocytic cells; therefore, trophoblast HLA-G may modulate the function of these cells. Co-culture with HLA-G(+) cells does not inhibit monocyte-derived dendritic cell up-regulation of HLA-DR and costimulatory molecules on maturation, but did increase production of IL-6 and IL-10. Furthermore, proliferation of allogeneic lymphocytes was inhibited by HLA-G binding to LILRB1/2 on responding antigen-presenting cells (APC). As HLA-G is the only HLA-I molecule that forms beta(2)m-associated dimers with increased avidity for LILRB1, this interaction could represent a placental-specific signal to decidual APC. We suggest that the placenta is modulating maternal immune responses locally in the uterus through HLA-G, a trophoblast-specific, monomorphic signal present in almost every pregnancy. See accompanying commentary: (http://dx.doi.org/10.1002/eji.200737515).
Figures






Comment in
-
Immune modulation of HLA-G dimer in maternal-fetal interface.Eur J Immunol. 2007 Jul;37(7):1727-9. doi: 10.1002/eji.200737515. Eur J Immunol. 2007. PMID: 17587197 Free PMC article. Review.
References
-
- McMaster MT, Librach CL, Zhou Y, Lim KH, Janatpour MJ, DeMars R, Kovats S, et al. Human placental HLA-G expression is restricted to differentiated cytotrophoblasts. J. Immunol. 1995;154:3771–3778. - PubMed
-
- Loke YW, King A, Burrows T, Gardner L, Bowen M, Hiby S, Howlett S, et al. Evaluation of trophoblast HLA-G antigen with a specific monoclonal antibody. Tissue Antigens. 1997;50:135–146. - PubMed
-
- Hiby SE, King A, Sharkey AM, Loke YW. Molecular studies of trophoblast HLA-G: Polymorphism, isoforms, imprinting and expression in preimplantation embryo. Tissue Antigens. 1999;53:1–13. - PubMed
-
- Blaschitz A, Juch H, Volz A, Hutter H, Daxboeck C, Desoye G, Dohr G. The soluble pool of HLA-G produced by human trophoblasts does not include detectable levels of the intron 4-containing HLA-G5 and HLA-G6 isoforms. Mol. Hum. Reprod. 2005;11:699–710. - PubMed
-
- Davis DM, Reyburn HT, Pazmany L, Chiu I, Mandelboim O, Strominger JL. Impaired spontaneous endocytosis of HLA-G. Eur. J. Immunol. 1997;27:2714–2719. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
