

Interleukin-13-induced type II polarization of inflammatory macrophages is mediated through suppression of nuclear factor-kappaB and preservation of IkappaBalpha in a T cell lymphoma
- PMID: 17550372
- PMCID: PMC1941949
- DOI: 10.1111/j.1365-2249.2007.03427.x
Interleukin-13-induced type II polarization of inflammatory macrophages is mediated through suppression of nuclear factor-kappaB and preservation of IkappaBalpha in a T cell lymphoma
Retraction in
-
Retraction statement.Clin Exp Immunol. 2009 Dec;158(3):334. doi: 10.1111/j.1365-2249.2009.04061.x. Clin Exp Immunol. 2009. PMID: 19906100 Free PMC article. No abstract available.
Abstract
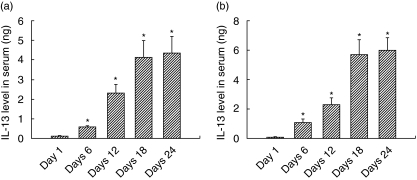
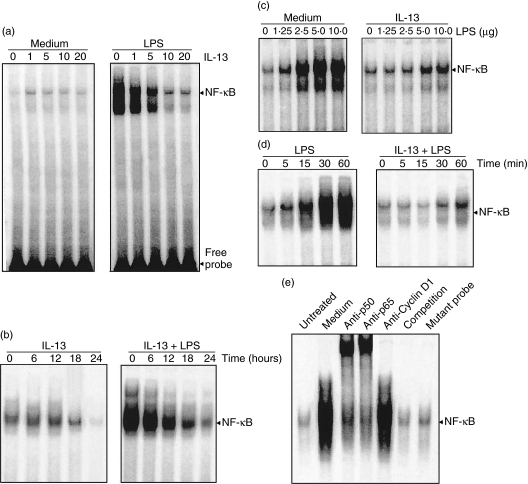
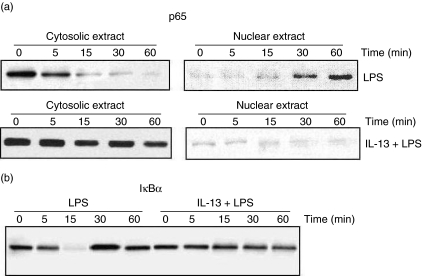
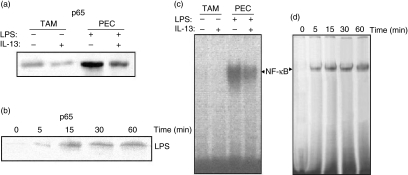
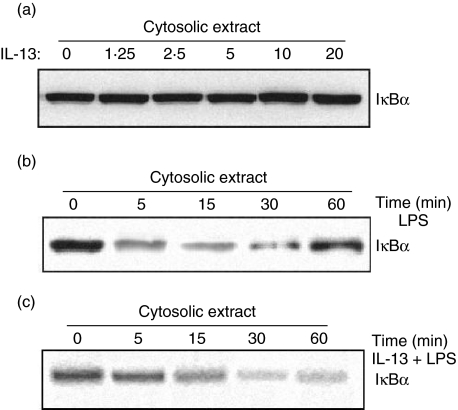
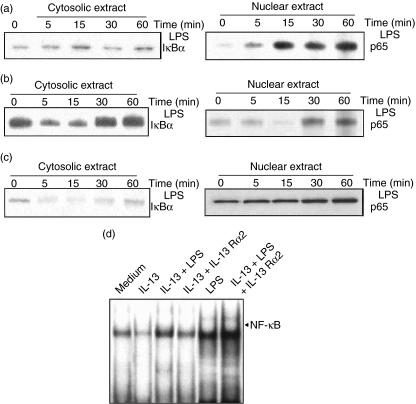
Spontaneously arising transplantable T cell lymphoma, designated as Dalton's lymphoma (DL), is characterized by a highly invasive and deleterious nature almost completely paralysing the host immune system. The level of interleukin (IL)-13 is elevated in serum and ascitic fluid of the DL-bearing host. IL-13 is a potent immunosuppressive cytokine and is an alternative activator of macrophages that suppresses the production of nitric oxide (NO) and expression of inducible nitric oxide synthase (iNOS), and proinflammatory cytokines. The expression of iNOS and proinflammatory cytokines are dependent largely upon the activation of nuclear factor-kappaB (NF-kappaB). Activation of NF-kappaB involves the degradation of cytoplasmic inhibitor IkappaBalpha, allowing the nuclear translocation of NF-kappaB and thereby transcription of the iNOS gene. Therefore, in this study we sought to determine whether the alternative activation or type II polarization of macrophages induced by IL-13 is mediated by the suppression of NF-kappaB and cytoplasmic preservation of IkappaBalpha. Western blot analysis and electrophoretic mobility shift assay (EMSA) indicate that tumour-associated macrophages (TAM) or polarized type II macrophages are due to preserved protein expression of IkappaBalpha, and therefore suppressed NF-kappaB nuclear translocation. These findings suggest that IL-13 may operate through the suppression of NF-kappaB activation and preservation of IkappaBalpha.
Figures
References
-
- Klein G. Comparative studies of mouse tumors with respect to their capacity for growth as ‘ascitic tumor’ and their average nucleic acid content. Exp Cell Res. 1951;2:518–24.
-
- Goldie H, Felix MD. Growth characteristics of free tumor cells transformed serially in the peritoneal fluid of mouse. Cancer Res. 1951;1:173–80. - PubMed
-
- Parajuli P, Singh SM, Kumar A, Sodhi A. Alterations in the tumoricidal function of murine tumor-associated macrophages during progressive growth of a tumor in vivo. Cancer J. 1997;10:222–7.
-
- Singh MP, Rai AK, Singh SM. Gender dimorphism in the progressive in vivo growth of a T-cell lymphoma: involvement of cytokines and gonadal hormones. J Reprod Immunol. 2005;65:17–32. - PubMed
-
- Minty A, Chalon P, Derocq J-M, et al. Interleukin-13 is a new human lymphokine regulating inflammatory and immune response. Nature. 1993;362:248–50. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
