

Slit-Robo interactions during cortical development
- PMID: 17553100
- PMCID: PMC2375773
- DOI: 10.1111/j.1469-7580.2007.00750.x
Slit-Robo interactions during cortical development
Abstract
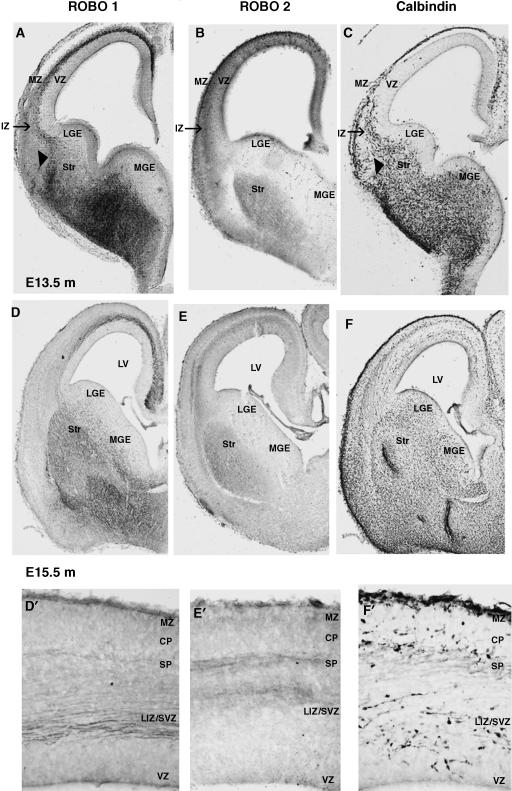
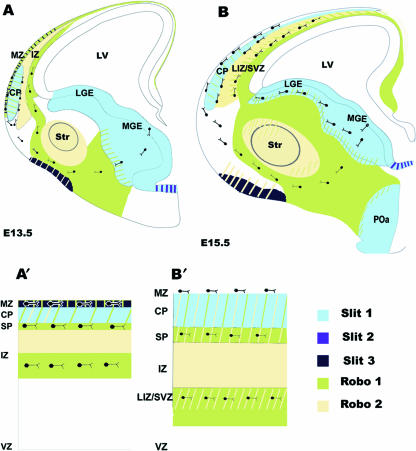
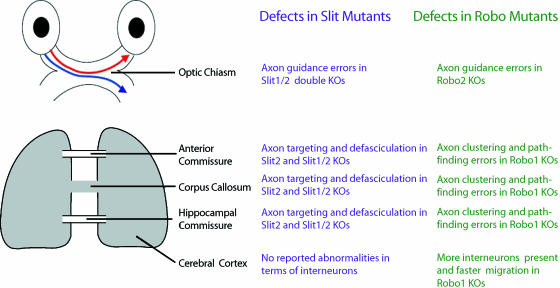
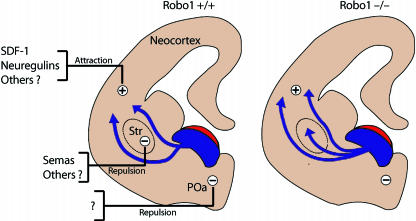
Interneurons are an integral part of cortical neuronal circuits. During the past decade, numerous studies have shown that these cells, unlike their pyramidal counterparts that are derived from the neuroepithelium along the lumen of the lateral ventricles, are generated in the ganglionic eminences in the subpallium. They use tangential migratory paths to reach the cortex, guided by intrinsic and extrinsic cues. Evidence is now emerging which suggests that the family of Slit proteins, acting through Robo receptors, play a role not only in axon guidance in the developing forebrain, but also as guiding signals in the migration of cortical interneurons. Here we describe the patterns of expression of Slit and Robo at different stages of forebrain development and review the evidence in support of their role in cortical interneuron migration. Slit-Robo signal transduction mechanisms are also important during normal development in a number of systems in the body and in disease states, making them potential therapeutic targets for the treatment of neurological disorders and certain types of cancer.
Figures
References
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Andrews W, Liapi A, Plachez A. Robo1 regulates the development of major axon tracts and interneuron migration in the forebrain. Development. 2006;133:2243–2252. - PubMed
-
- Bagri A, Marín O, Plump AS. Slit prevents midline crossing and determines the dorsoventral position of major axonal pathways in the mammalian forebrain. Neuron. 2002;33:233–248. - PubMed
-
- Bashaw GJ, Kidd T, Murray D, Pawson T, Goodman CS. Repulsion axon guidance: Abelson and Enabled play opposing roles downstream of the roundabout receptor. Cell. 2000;101:703–715. - PubMed
-
- Battye R, Stevens A, Jacobs JR. Axon repulsion from the midline of the Drosophila CNS requires slit function. Development. 1999;126:2475–2481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
