

Neuronal mechanism for neuropathic pain
- PMID: 17553143
- PMCID: PMC1904186
- DOI: 10.1186/1744-8069-3-14
Neuronal mechanism for neuropathic pain
Abstract
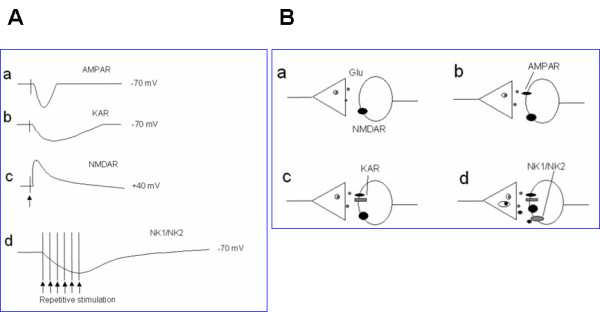
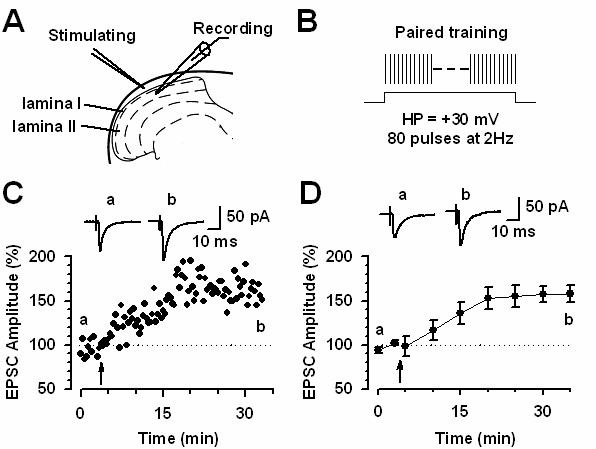
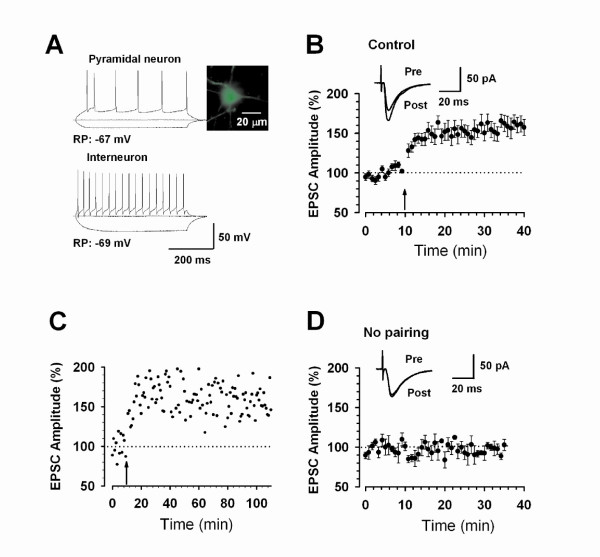
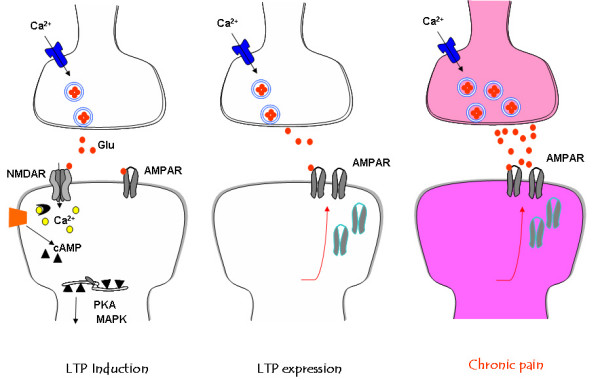
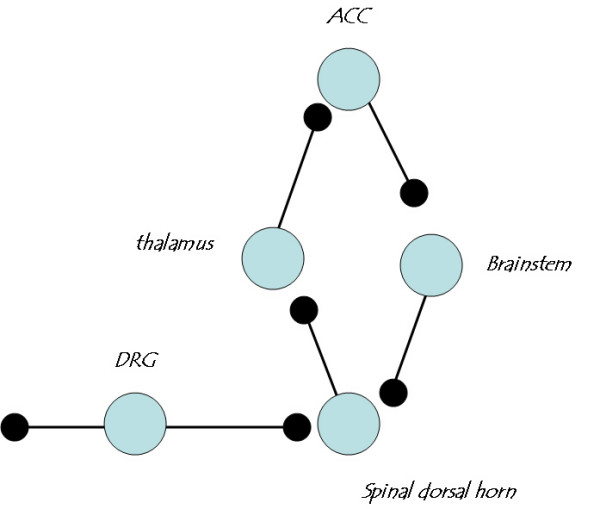
Among different forms of persistent pain, neuropathic pain presents as a most difficult task for basic researchers and clinicians. Despite recent rapid development of neuroscience and modern techniques related to drug discovery, effective drugs based on clear basic mechanisms are still lacking. Here, I will review the basic neuronal mechanisms that maybe involved in neuropathic pain. I will present the problem of neuropathic pain as a rather difficult task for neuroscientists, and we may have to wait for a long time before we fully understand how brain encode, store, and retrieve painful information after the injury. I propose that neuropathic pain as a major brain disease, rather being a clinic problem due to peripheral injury.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
