

Thermal limits and adaptation in marine Antarctic ectotherms: an integrative view
- PMID: 17553776
- PMCID: PMC2443174
- DOI: 10.1098/rstb.2006.1947
Thermal limits and adaptation in marine Antarctic ectotherms: an integrative view
Abstract
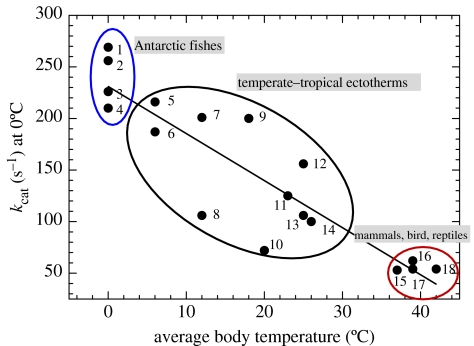
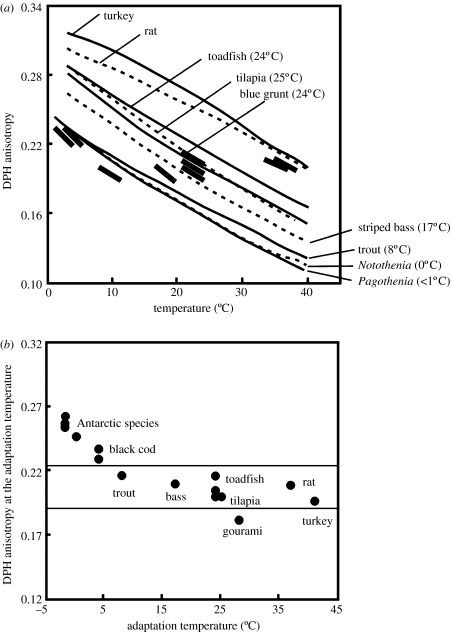
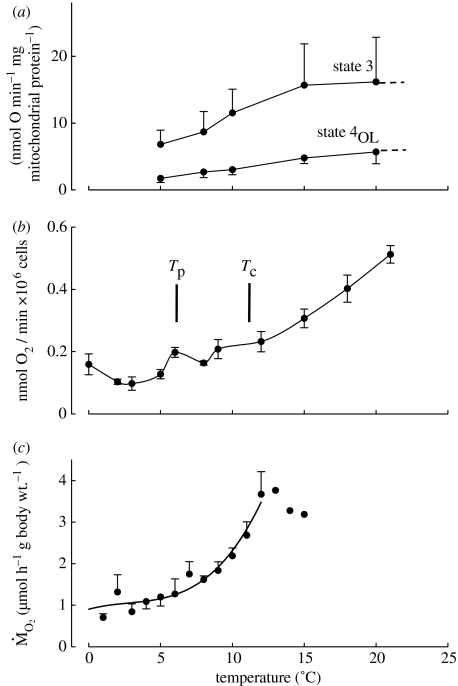
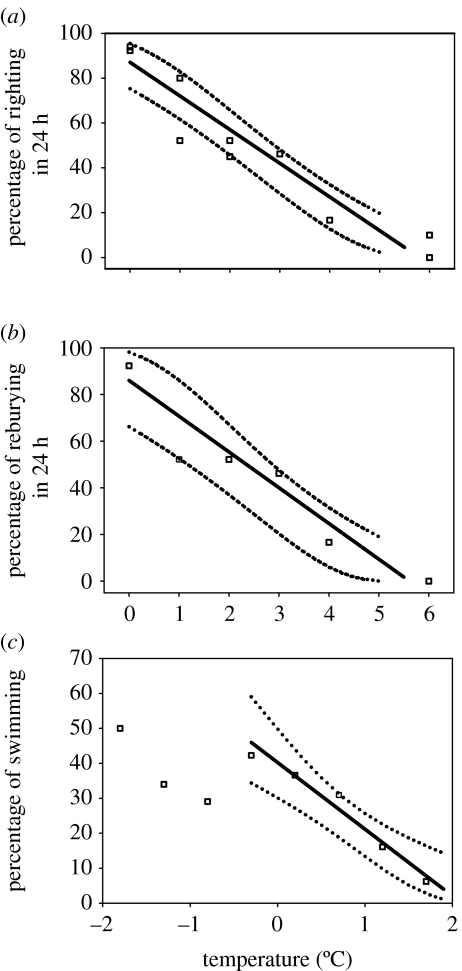
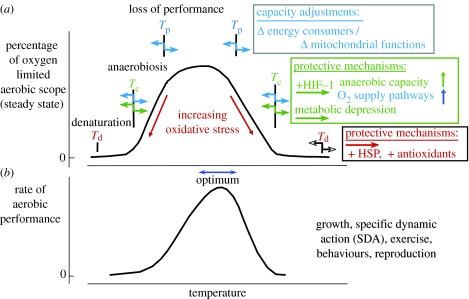
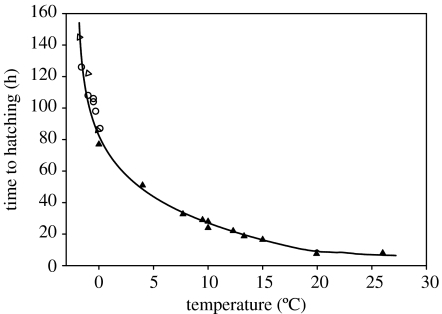
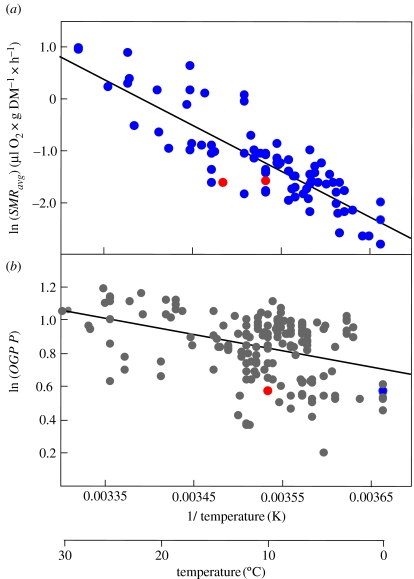
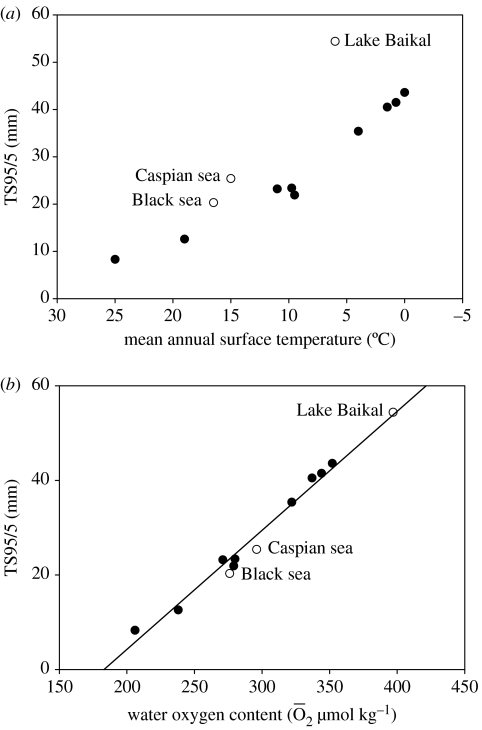
A cause and effect understanding of thermal limitation and adaptation at various levels of biological organization is crucial in the elaboration of how the Antarctic climate has shaped the functional properties of extant Antarctic fauna. At the same time, this understanding requires an integrative view of how the various levels of biological organization may be intertwined. At all levels analysed, the functional specialization to permanently low temperatures implies reduced tolerance of high temperatures, as a trade-off. Maintenance of membrane fluidity, enzyme kinetic properties (Km and k(cat)) and protein structural flexibility in the cold supports metabolic flux and regulation as well as cellular functioning overall. Gene expression patterns and, even more so, loss of genetic information, especially for myoglobin (Mb) and haemoglobin (Hb) in notothenioid fishes, reflect the specialization of Antarctic organisms to a narrow range of low temperatures. The loss of Mb and Hb in icefish, together with enhanced lipid membrane densities (e.g. higher concentrations of mitochondria), becomes explicable by the exploitation of high oxygen solubility at low metabolic rates in the cold, where an enhanced fraction of oxygen supply occurs through diffusive oxygen flux. Conversely, limited oxygen supply to tissues upon warming is an early cause of functional limitation. Low standard metabolic rates may be linked to extreme stenothermy. The evolutionary forces causing low metabolic rates as a uniform character of life in Antarctic ectothermal animals may be linked to the requirement for high energetic efficiency as required to support higher organismic functioning in the cold. This requirement may result from partial compensation for the thermal limitation of growth, while other functions like hatching, development, reproduction and ageing are largely delayed. As a perspective, the integrative approach suggests that the patterns of oxygen- and capacity-limited thermal tolerance are linked, on one hand, with the capacity and design of molecules and membranes, and, on the other hand, with life-history consequences and lifestyles typically seen in the permanent cold. Future research needs to address the detailed aspects of these interrelationships.
Figures
References
-
- Abele D, Burlando B, Viarengo A, Pörtner H.O. Exposure to elevated temperatures and hydrogen peroxide elicits oxidative stress and antioxidant response in the Antarctic intertidal limpet Nacella concinna. Comp. Biochem. Physiol. B. 1998;120:425–435.
-
- Acierno R, Agnisola C, Tota B, Sidell B.D. Myoglobin enhances cardiac performance in Antarctic fish species that express the protein. Am. J. Physiol. 1997;273:R100–R106. - PubMed
-
- Aizawa A.S, Nishino H, Saito K, Kimura K, Shirakawa H, Yoshida M. Stimulation of transcription in cultured cells by high mobility group protein 1: essential role of the acidic carboxyl-terminal region. Biochemistry. 1994;33:14 690–14 695. - PubMed
-
- Angel M.V. Variations in time and space: is biogeography relevant to studies of long-time scale change? J. Mar. Biol. Assoc. UK. 1991;71:191–206.
-
- Arntz W.E, Brey T, Gallardo V.A. Antarctic zoobenthos. Oceangr. Mar. Biol.: Annu. Rev. 1994;32:241–304.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous