

Elastic properties of ribosomal RNA building blocks: molecular dynamics of the GTPase-associated center rRNA
- PMID: 17553840
- PMCID: PMC1919483
- DOI: 10.1093/nar/gkm245
Elastic properties of ribosomal RNA building blocks: molecular dynamics of the GTPase-associated center rRNA
Abstract
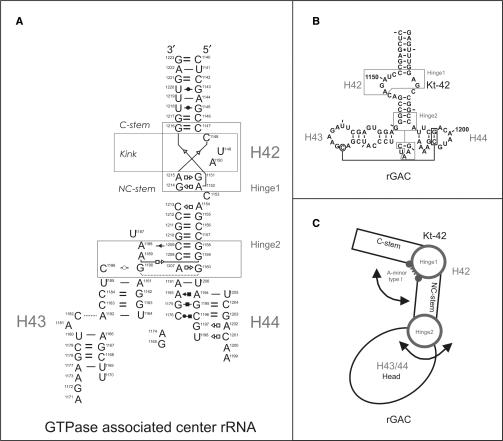
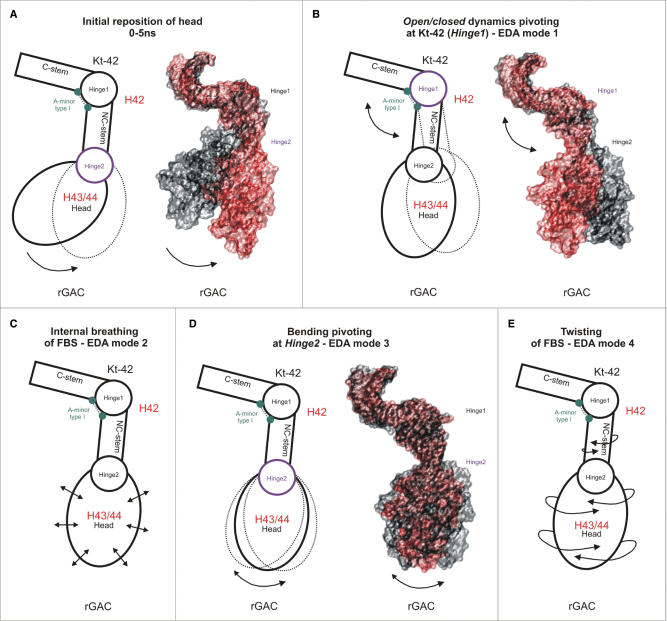
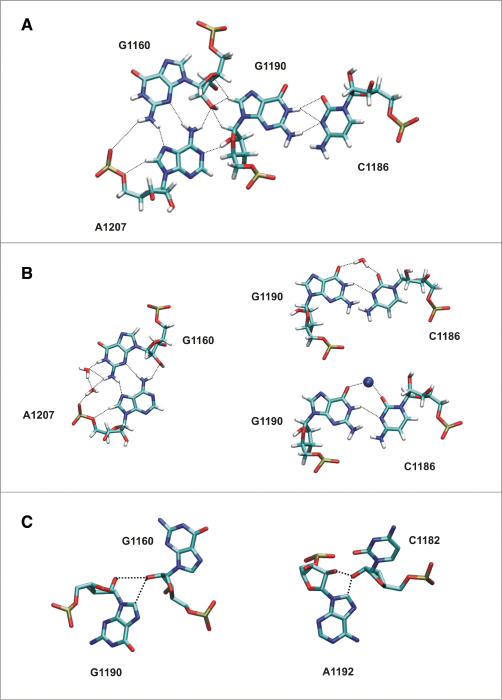
Explicit solvent molecular dynamics (MD) was used to describe the intrinsic flexibility of the helix 42-44 portion of the 23S rRNA (abbreviated as Kt-42+rGAC; kink-turn 42 and GTPase-associated center rRNA). The bottom part of this molecule consists of alternating rigid and flexible segments. The first flexible segment (Hinge1) is the highly anharmonic kink of Kt-42. The second one (Hinge2) is localized at the junction between helix 42 and helices 43/44. The rigid segments are the two arms of helix 42 flanking the kink. The whole molecule ends up with compact helices 43/44 (Head) which appear to be modestly compressed towards the subunit in the Haloarcula marismortui X-ray structure. Overall, the helix 42-44 rRNA is constructed as a sophisticated intrinsically flexible anisotropic molecular limb. The leading flexibility modes include bending at the hinges and twisting. The Head shows visible internal conformational plasticity, stemming from an intricate set of base pairing patterns including dynamical triads and tetrads. In summary, we demonstrate how rRNA building blocks with contrasting intrinsic flexibilities can form larger architectures with highly specific patterns of preferred low-energy motions and geometries.
Figures
Similar articles
-
Molecular dynamics simulations suggest that RNA three-way junctions can act as flexible RNA structural elements in the ribosome.Nucleic Acids Res. 2010 Oct;38(18):6247-64. doi: 10.1093/nar/gkq414. Epub 2010 May 27. Nucleic Acids Res. 2010. PMID: 20507916 Free PMC article.
-
Ribosomal RNA kink-turn motif--a flexible molecular hinge.J Biomol Struct Dyn. 2004 Oct;22(2):183-94. doi: 10.1080/07391102.2004.10506994. J Biomol Struct Dyn. 2004. PMID: 15317479
-
The kink-turn: a new RNA secondary structure motif.EMBO J. 2001 Aug 1;20(15):4214-21. doi: 10.1093/emboj/20.15.4214. EMBO J. 2001. PMID: 11483524 Free PMC article.
-
Hinge-like motions in RNA kink-turns: the role of the second a-minor motif and nominally unpaired bases.Biophys J. 2005 May;88(5):3466-85. doi: 10.1529/biophysj.104.054916. Epub 2005 Feb 18. Biophys J. 2005. PMID: 15722438 Free PMC article.
-
The archaeon Haloarcula marismortui has few modifications in the central parts of its 23S ribosomal RNA.J Mol Biol. 2005 May 6;348(3):563-73. doi: 10.1016/j.jmb.2005.03.009. J Mol Biol. 2005. PMID: 15826654
Cited by
-
Connecting the kinetics and energy landscape of tRNA translocation on the ribosome.PLoS Comput Biol. 2013;9(3):e1003003. doi: 10.1371/journal.pcbi.1003003. Epub 2013 Mar 21. PLoS Comput Biol. 2013. PMID: 23555233 Free PMC article.
-
Dynamics of the base of ribosomal A-site finger revealed by molecular dynamics simulations and Cryo-EM.Nucleic Acids Res. 2010 Mar;38(4):1325-40. doi: 10.1093/nar/gkp1057. Epub 2009 Dec 1. Nucleic Acids Res. 2010. PMID: 19952067 Free PMC article.
-
RNA Structural Dynamics As Captured by Molecular Simulations: A Comprehensive Overview.Chem Rev. 2018 Apr 25;118(8):4177-4338. doi: 10.1021/acs.chemrev.7b00427. Epub 2018 Jan 3. Chem Rev. 2018. PMID: 29297679 Free PMC article. Review.
-
Molecular dynamics simulations suggest that RNA three-way junctions can act as flexible RNA structural elements in the ribosome.Nucleic Acids Res. 2010 Oct;38(18):6247-64. doi: 10.1093/nar/gkq414. Epub 2010 May 27. Nucleic Acids Res. 2010. PMID: 20507916 Free PMC article.
-
An RNA molecular switch: Intrinsic flexibility of 23S rRNA Helices 40 and 68 5'-UAA/5'-GAN internal loops studied by molecular dynamics methods.J Chem Theory Comput. 2010 Jan 1;2010(6):910-929. J Chem Theory Comput. 2010. PMID: 21132104 Free PMC article.
References
-
- Ramakrishnan V. Ribosome structure and the mechanism of translation. Cell. 2002;108:557–572. - PubMed
-
- Moore PB, Steitz TA. The structural basis of large ribosomal subunit function. Annu. Rev. Biochem. 2003;72:813–850. - PubMed
-
- Frank J, Agrawal RK. A ratchet-like inter-subunit reorganization of the ribosome during translocation. Nature. 2000;406:318–322. - PubMed
-
- Wimberly BT, Brodersen DE, Clemons WM, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan V. Structure of the 30S ribosomal subunit. Nature. 2000;407:327–339. - PubMed
-
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA. The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science. 2000;289:905–920. - PubMed
