

E-cadherin adhesion activates c-Src signaling at cell-cell contacts
- PMID: 17553930
- PMCID: PMC1949350
- DOI: 10.1091/mbc.e06-12-1154
E-cadherin adhesion activates c-Src signaling at cell-cell contacts
Abstract
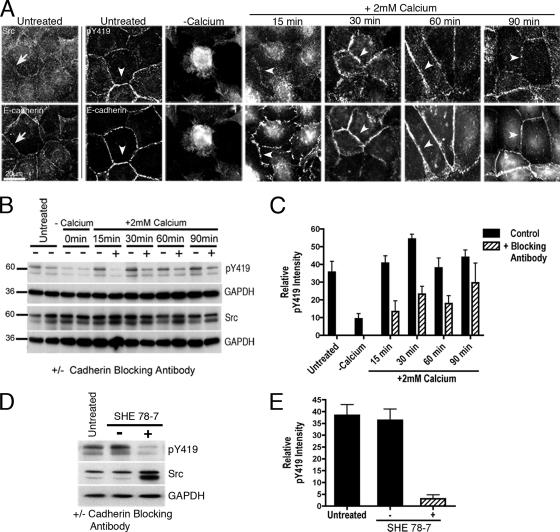
Cadherin-based cell-cell contacts are prominent sites for phosphotyrosine signaling, being enriched in tyrosine-phosphorylated proteins and tyrosine kinases and phosphatases. The functional interplay between cadherin adhesion and tyrosine kinase signaling, however, is complex and incompletely understood. In this report we tested the hypothesis that cadherin adhesion activates c-Src signaling and sought to assess its impact on cadherin function. We identified c-Src as part of a cadherin-activated cell signaling pathway that is stimulated by ligation of the adhesion receptor. However, c-Src has a biphasic impact on cadherin function, exerting a positive supportive role at lower signal strengths, but inhibiting function at high signal strengths. Inhibiting c-Src under circumstances when it is activated by cadherin adhesion decreased several measures of cadherin function. This suggests that the cadherin-activated c-Src signaling pathway serves positively to support cadherin function. Finally, our data implicate PI3-kinase signaling as a target for cadherin-activated c-Src signaling that contributes to its positive impact on cadherin function. We conclude that E-cadherin signaling is an important activator of c-Src at cell-cell contacts, providing a key input into a signaling pathway where quantitative changes in signal strength may result in qualitative differences in functional outcome.
Figures







Similar articles
-
Recruitment of phosphoinositide 3-kinase defines a positive contribution of tyrosine kinase signaling to E-cadherin function.J Biol Chem. 2005 Jan 28;280(4):3043-50. doi: 10.1074/jbc.M412148200. Epub 2004 Nov 19. J Biol Chem. 2005. PMID: 15556934
-
The catalytic activity of the Src family kinases is required to disrupt cadherin-dependent cell-cell contacts.Mol Biol Cell. 2000 Jan;11(1):51-64. doi: 10.1091/mbc.11.1.51. Mol Biol Cell. 2000. PMID: 10637290 Free PMC article.
-
E-cadherin homophilic ligation directly signals through Rac and phosphatidylinositol 3-kinase to regulate adhesive contacts.J Biol Chem. 2002 Feb 22;277(8):6708-18. doi: 10.1074/jbc.M109640200. Epub 2001 Dec 13. J Biol Chem. 2002. PMID: 11744701
-
The SRC-induced mesenchymal state in late-stage colon cancer cells.Cells Tissues Organs. 2005;179(1-2):73-80. doi: 10.1159/000084511. Cells Tissues Organs. 2005. PMID: 15942195 Review.
-
Not so simple: the complexity of phosphotyrosine signaling at cadherin adhesive contacts.J Mol Med (Berl). 2007 Jun;85(6):545-54. doi: 10.1007/s00109-007-0198-x. Epub 2007 Apr 11. J Mol Med (Berl). 2007. PMID: 17429596 Review.
Cited by
-
The claudin-transcription factor signaling pathway.Tissue Barriers. 2021 Jul 3;9(3):1908109. doi: 10.1080/21688370.2021.1908109. Epub 2021 Apr 27. Tissue Barriers. 2021. PMID: 33906582 Free PMC article. Review.
-
Involvement of Src in the membrane skeletal complex, MPP6-4.1G, in Schmidt-Lanterman incisures of mouse myelinated nerve fibers in PNS.Histochem Cell Biol. 2013 Aug;140(2):213-22. doi: 10.1007/s00418-012-1073-6. Epub 2013 Jan 10. Histochem Cell Biol. 2013. PMID: 23306908
-
Indometacin ameliorates high glucose-induced proliferation and invasion via modulation of e-cadherin in pancreatic cancer cells.Curr Med Chem. 2013;20(33):4142-4152. doi: 10.2174/09298673113209990249. Curr Med Chem. 2013. PMID: 23992308 Free PMC article.
-
Recruitment of Lyn from endomembranes to the plasma membrane through calcium-dependent cell-cell interactions upon polarization of inducible Lyn-expressing MDCK cells.Sci Rep. 2017 Mar 28;7(1):493. doi: 10.1038/s41598-017-00538-5. Sci Rep. 2017. PMID: 28352128 Free PMC article.
-
Adherens Junctions on the Move-Membrane Trafficking of E-Cadherin.Cold Spring Harb Perspect Biol. 2017 Mar 1;9(3):a029140. doi: 10.1101/cshperspect.a029140. Cold Spring Harb Perspect Biol. 2017. PMID: 28096264 Free PMC article. Review.
References
-
- Avizienyte E., Wyke A. W., Jones R. J., McLean G. W., Westhoff M. A., Brunton V. G., Frame M. C. Src-induced de-regulation of E-cadherin in colon cancer cells requires integrin signalling. Nat. Cell Biol. 2002;4:632–638. - PubMed
-
- Behrens J., Vakaet L., Friis R., Winterhager E., Van Roy F., Mareel M. M., Birchmeier W. Loss of epithelial differentiation and gain of invasiveness correlates with tyrosine phosphorylation of the E-cadherin/beta-catenin complex in cells transformed with a temperature-sensitive v-SRC gene. J. Cell Biol. 1993;120:757–766. - PMC - PubMed
-
- Cantrell D. A. Phosphoinositide 3-kinase signalling pathways. J. Cell Sci. 2001;114:1439–1445. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
