

Human p53 is inhibited by glutathionylation of cysteines present in the proximal DNA-binding domain during oxidative stress
- PMID: 17555331
- PMCID: PMC2518322
- DOI: 10.1021/bi700425y
Human p53 is inhibited by glutathionylation of cysteines present in the proximal DNA-binding domain during oxidative stress
Abstract
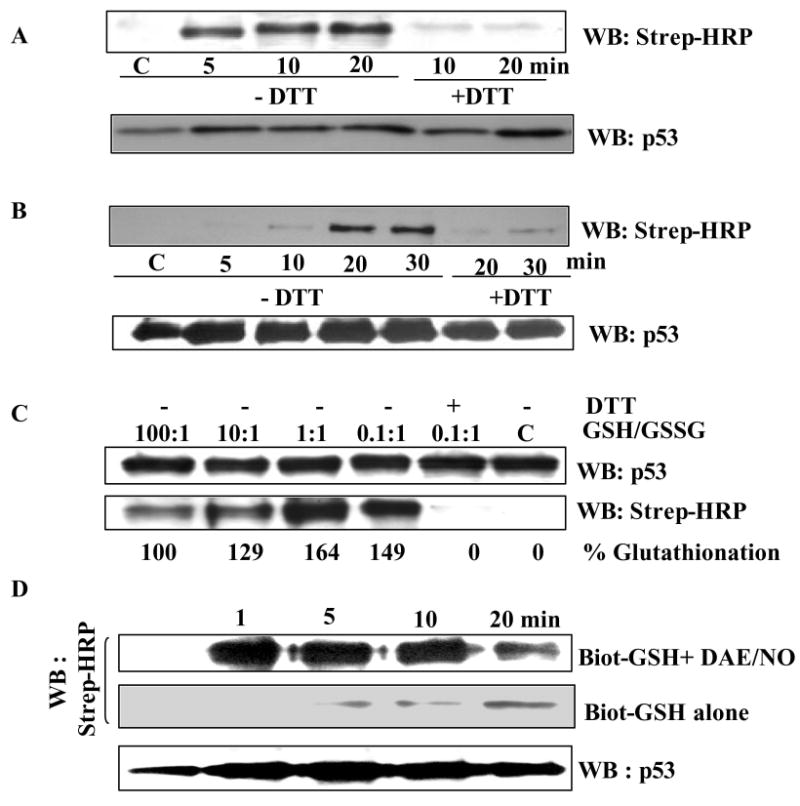
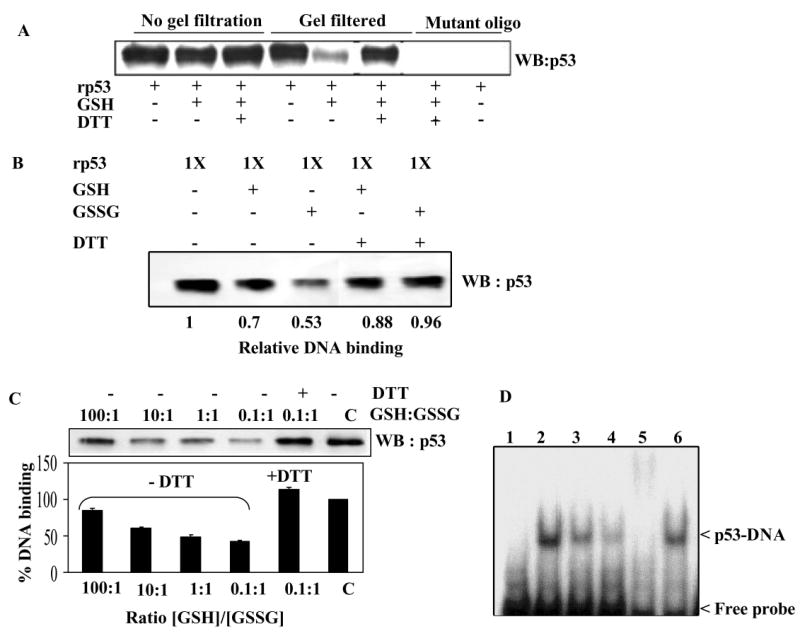
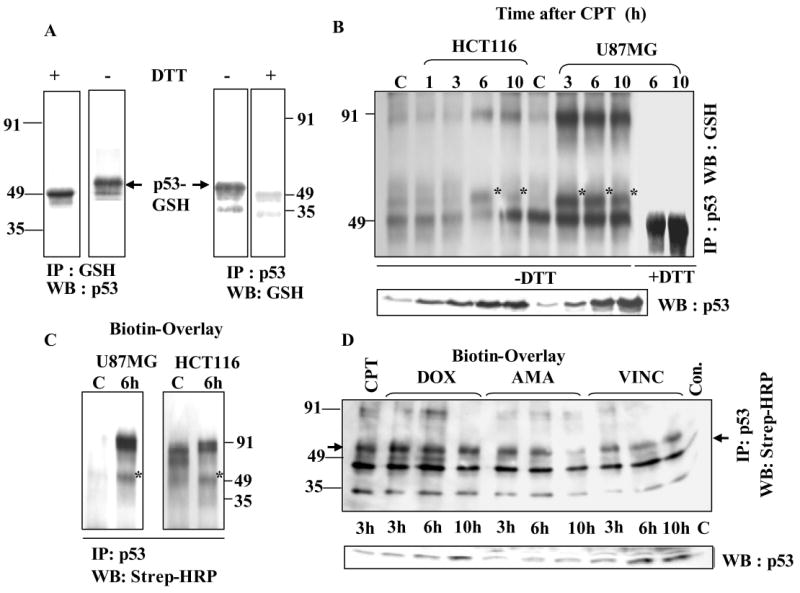
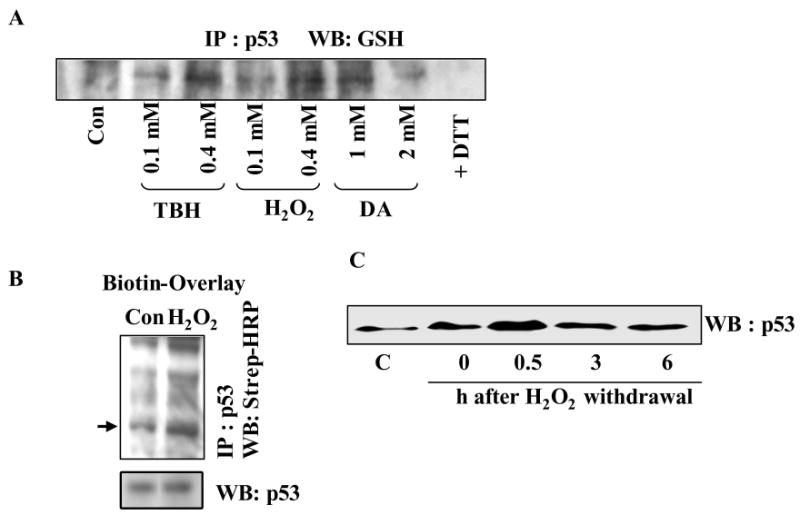
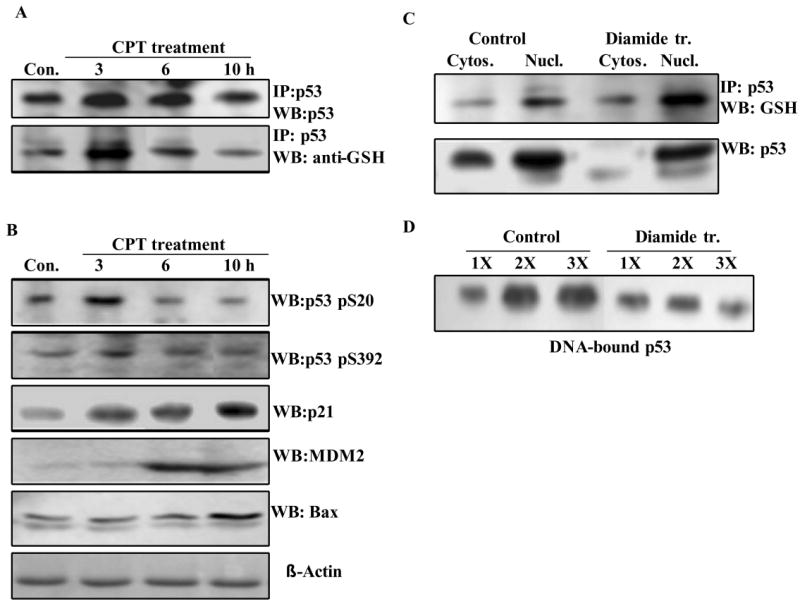
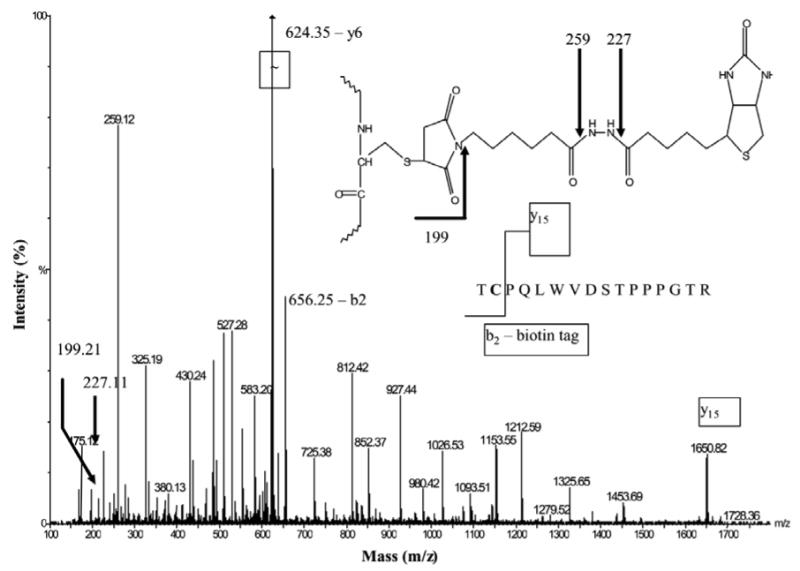
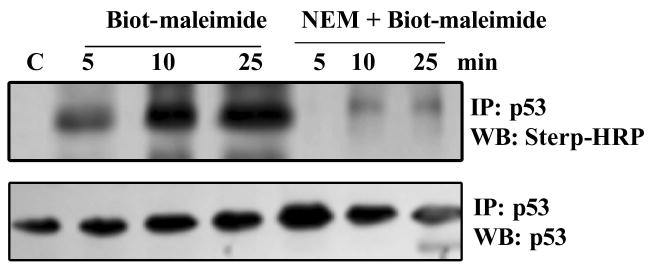
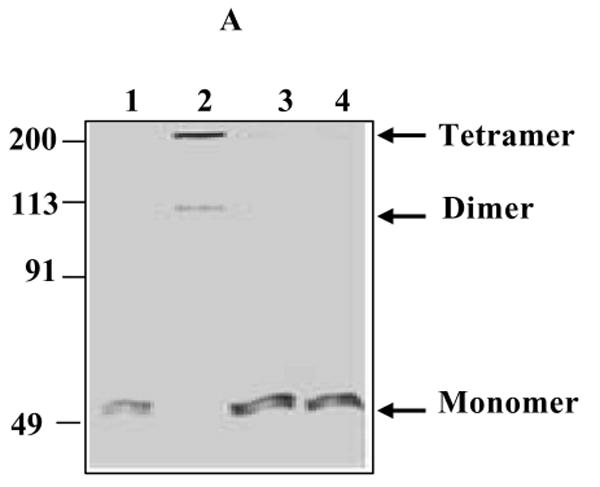
The cellular mechanisms that modulate the redox state of p53 tumor suppressor remain unclear, although its DNA binding function is known to be strongly inhibited by oxidative and nitrosative stresses. We show that human p53 is subjected to a new and reversible posttranslational modification, namely, S-glutathionylation in stressed states, including DNA damage. First, a rapid and direct incorporation of biotinylated GSH or GSSG into the purified recombinant p53 protein was observed. The modified p53 had a significantly weakened ability to bind its consensus DNA sequence. Reciprocal immunoprecipitations and a GST overlay assay showed that p53 in tumor cells was marginally glutathionylated; however, the level of modification increased greatly after oxidant and DNA-damaging treatments. GSH modification coexisted with the serine phophorylations in activated p53, and the thiol-conjugated protein was present in nuclei. When tumor cells treated with camptothecin or cisplatin were subsequently exposed to glutathione-enhancing agents, p53 underwent dethiolation accompanied by detectable increases in the level of p21waf1 expression, relative to the DNA-damaging drugs alone. Mass spectrometry of GSH-modified p53 protein identified cysteines 124, 141, and 182, all present in the proximal DNA-binding domain, as the sites of glutathionylation. Biotinylated maleimide also reacted rapidly with Cys141, implying that this is the most reactive cysteine on the p53 surface. The glutathionylatable cysteines were found to exist in a negatively charged microenvironment in cellular p53. Molecular modeling studies located Cys124 and -141 at the dimer interface of p53 and showed glutathionylation of either residue would inhibit p53-DNA association and also interfere with protein dimerization. These results show for the first time that shielding of reactive cysteines contributes to a negative regulation for human p53 and imply that such an inactivation of the transcription factor may represent an acute defensive response with significant consequences for oncogenesis.
Figures


References
-
- Mills AA. p53: link to the past bridge to the future. Genes Develop. 2005;19:2091–2099. - PubMed
-
- Inoue T, Wu L, Stuart J, Maki CG. Control of p53 nuclear accumulation in stressed cells. FEBS Lett. 2005;579:4978–4984. - PubMed
-
- Strosznajder RP, Jesko H, Banasik M, Tanaka S. Effects of p53 inhibitor on survival and death of cells subjected to oxidative stress. J Physiol Pharmacol. 2005;4:215–221. - PubMed
-
- Bode AM, Dong Z. Post-translational modification of p53 in tumorigenesis. Nat Rev Cancer. 2004;4:793–805. - PubMed
-
- Martindale JL, Holbrook NJ. Cellular response to oxidative stress: signaling for suicide and survival. J Cellular Physiol. 2002;192:1–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
