

CD19 expression in B cells is important for suppression of contact hypersensitivity
- PMID: 17556590
- PMCID: PMC1934538
- DOI: 10.2353/ajpath.2007.061279
CD19 expression in B cells is important for suppression of contact hypersensitivity
Abstract
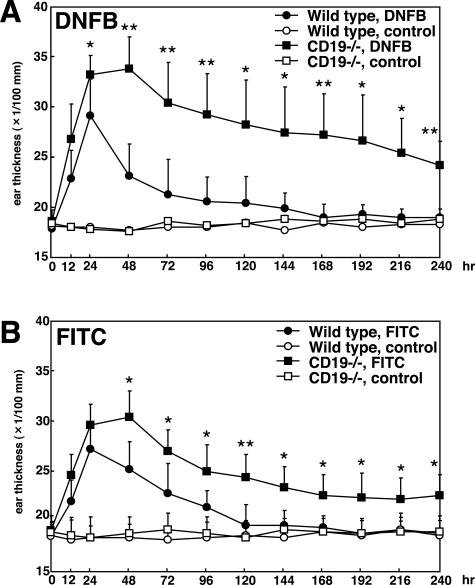
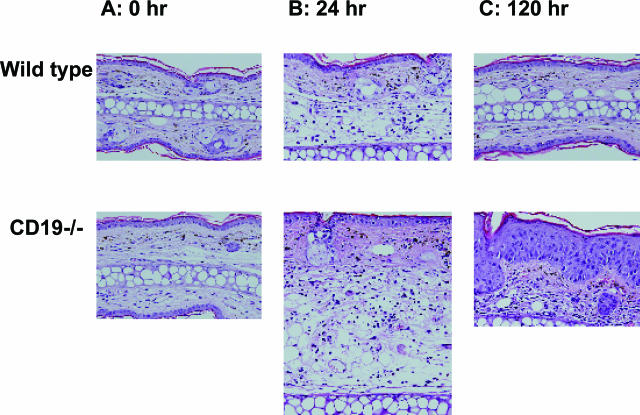
Contact hypersensitivity (CHS) is a cutaneous immune reaction mediated mainly by antigen-specific effector T cells and is regarded as a model for Th1/Tc1-mediated inflammation. However, recent reports have suggested pivotal roles of B cells in CHS. CD19 serves as a positive B-cell response regulator that defines signaling thresholds critical for B-cell responses. In the current study, we assessed the role of the B-cell-specific surface molecule CD19 on the development of CHS by examining CD19-deficient mice. Although CD19-deficient mice are hyposensitive to a variety of transmembrane signals, CD19 loss resulted in increased and prolonged reaction of CHS, suggesting an inhibitory role of CD19 expression in CHS. Sensitized lymph nodes and elicited ear lesions from CD19-deficient mice exhibited Th1/Tc1-shifted cytokine profile with increased interferon-gamma expression and decreased interleukin-10 expression. Adoptive transfer experiments revealed that CD19 expression in recipient mice was required for optimal suppression of CHS response, indicating its role in the elicitation phase. Furthermore, spleen B cells, especially marginal zone B cells, from wild-type mice were able to normalize exaggerated CHS reactions in CD19-deficient mice. Thus, CD19 expression in B cells is critical for termination of CHS responses, possibly through the function of regulatory B cells.
Figures








Similar articles
-
B cells are not required for T cell priming in low zone tolerance to contact allergens and contact hypersensitivity.Eur J Immunol. 2004 Nov;34(11):3082-90. doi: 10.1002/eji.200425402. Eur J Immunol. 2004. PMID: 15376190
-
The Role of Interleukin-19 in Contact Hypersensitivity.Biol Pharm Bull. 2018;41(2):182-189. doi: 10.1248/bpb.b17-00594. Biol Pharm Bull. 2018. PMID: 29386478
-
CD4+ Th1 and CD8+ type 1 cytotoxic T cells both play a crucial role in the full development of contact hypersensitivity.J Immunol. 2000 Dec 15;165(12):6783-90. doi: 10.4049/jimmunol.165.12.6783. J Immunol. 2000. PMID: 11120799
-
Cytokine knockouts in contact hypersensitivity research.Cytokine Growth Factor Rev. 2003 Oct;14(5):381-9. doi: 10.1016/s1359-6101(03)00050-9. Cytokine Growth Factor Rev. 2003. PMID: 12948522 Review.
-
The role of NO in contact hypersensitivity.Int Immunopharmacol. 2001 Aug;1(8):1469-78. doi: 10.1016/s1567-5769(01)00091-1. Int Immunopharmacol. 2001. PMID: 11515812 Review.
Cited by
-
Distribution characteristics of circulating B cell subpopulations in patients with chronic kidney disease.Sci Rep. 2023 Nov 27;13(1):20797. doi: 10.1038/s41598-023-47742-0. Sci Rep. 2023. PMID: 38012211 Free PMC article.
-
Regulatory and Effector B Cells: A New Path Toward Biomarkers and Therapeutic Targets to Improve Transplant Outcomes?Clin Lab Med. 2019 Mar;39(1):15-29. doi: 10.1016/j.cll.2018.10.011. Epub 2018 Dec 18. Clin Lab Med. 2019. PMID: 30709503 Free PMC article. Review.
-
B-cell-mediated strategies to fight chronic allograft rejection.Front Immunol. 2013 Dec 17;4:444. doi: 10.3389/fimmu.2013.00444. Front Immunol. 2013. PMID: 24381571 Free PMC article. Review.
-
CD22 expression mediates the regulatory functions of peritoneal B-1a cells during the remission phase of contact hypersensitivity reactions.J Immunol. 2010 May 1;184(9):4637-45. doi: 10.4049/jimmunol.0901719. Epub 2010 Mar 24. J Immunol. 2010. PMID: 20335532 Free PMC article.
-
Impaired humoral immunity and tolerance in K14-VEGFR-3-Ig mice that lack dermal lymphatic drainage.J Immunol. 2012 Sep 1;189(5):2181-90. doi: 10.4049/jimmunol.1103545. Epub 2012 Jul 27. J Immunol. 2012. PMID: 22844119 Free PMC article.
References
-
- Gorbachev AV, Fairchild RL. Induction and regulation of T-cell priming for contact hypersensitivity. Crit Rev Immunol. 2001;21:451–472. - PubMed
-
- Wang B, Feliciani C, Howell BG, Freed I, Cai Q, Watanabe H, Sauder DN. Contribution of Langerhans cell-derived IL-18 to contact hypersensitivity. J Immunol. 2002;168:3303–3308. - PubMed
-
- Marchal G, Seman M, Milon G, Truffa-Bachi P, Zilberfarb V. Local adoptive transfer of skin delayed-type hypersensitivity initiated by a single T lymphocyte. J Immunol. 1982;129:954–958. - PubMed
-
- Kalish RS, Askenase PW. Molecular mechanisms of CD8+ T cell-mediated delayed hypersensitivity: implications for allergies, asthma, and autoimmunity. J Allergy Clin Immunol. 1999;103:192–199. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
