

Radiation and inhibition of angiogenesis by canstatin synergize to induce HIF-1alpha-mediated tumor apoptotic switch
- PMID: 17557121
- PMCID: PMC1884687
- DOI: 10.1172/JCI30269
Radiation and inhibition of angiogenesis by canstatin synergize to induce HIF-1alpha-mediated tumor apoptotic switch
Abstract
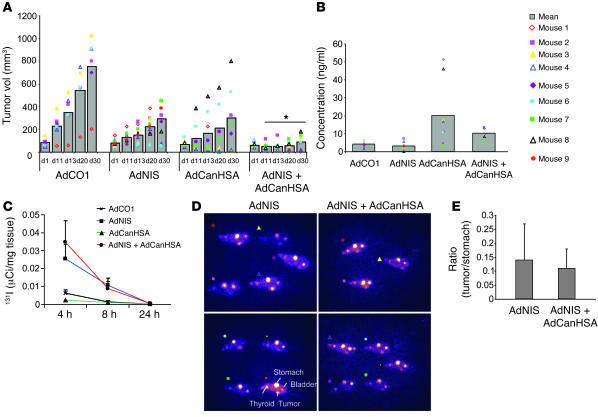
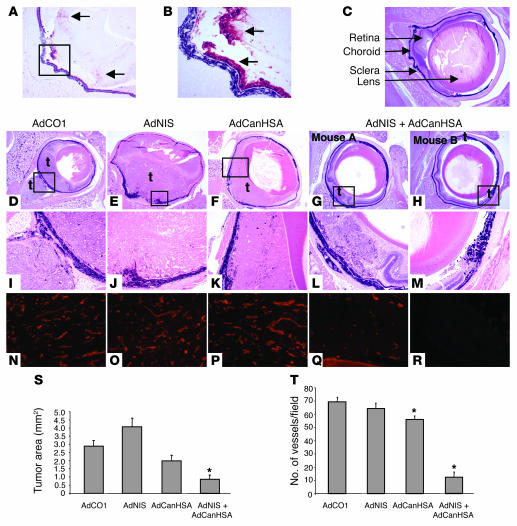
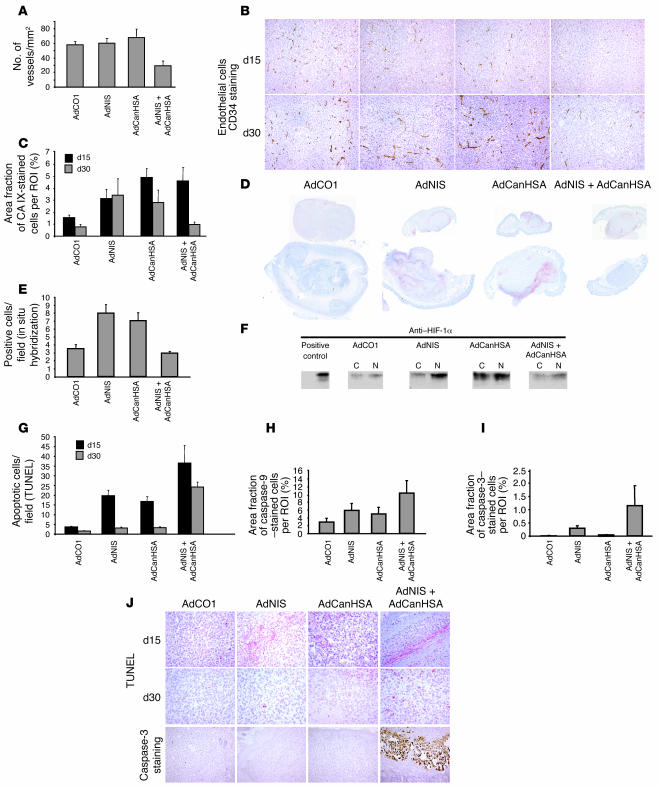
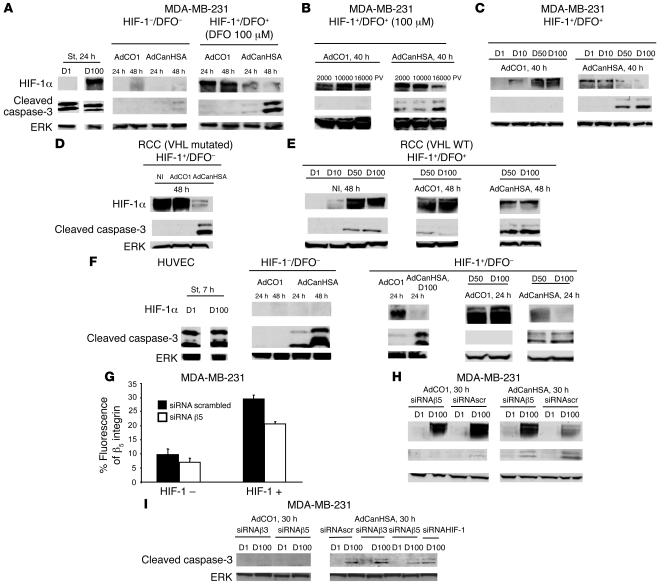
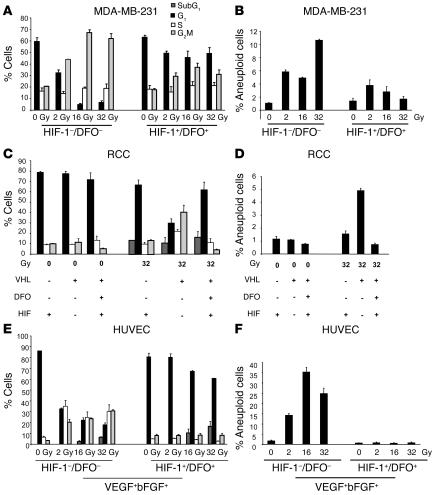
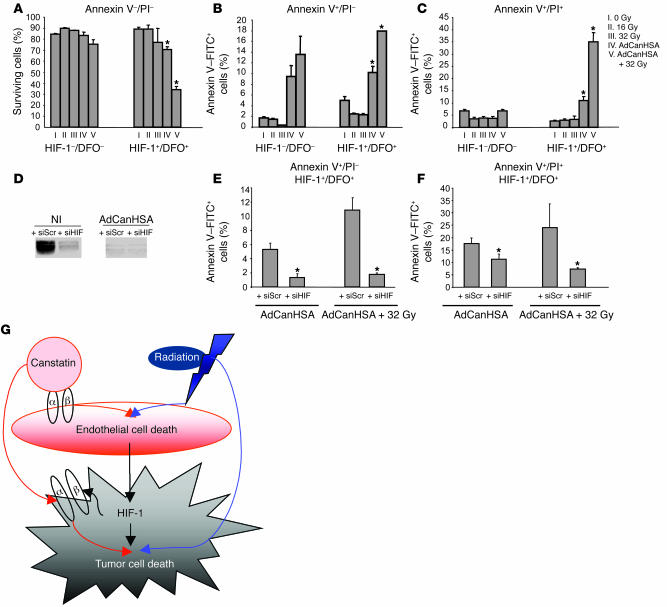
Tumor radioresponsiveness depends on endothelial cell death, which leads in turn to tumor hypoxia. Radiation-induced hypoxia was recently shown to trigger tumor radioresistance by activating angiogenesis through hypoxia-inducible factor 1-regulated (HIF-1-regulated) cytokines. We show here that combining targeted radioiodide therapy with angiogenic inhibitors, such as canstatin, enhances direct tumor cell apoptosis, thereby overcoming radio-induced HIF-1-dependent tumor survival pathways in vitro and in vivo. We found that following dual therapy, HIF-1alpha increases the activity of the canstatin-induced alpha(v)beta(5) signaling tumor apoptotic pathway and concomitantly abrogates mitotic checkpoint and tetraploidy triggered by radiation. Apoptosis in conjunction with mitotic catastrophe leads to lethal tumor damage. We discovered that HIF-1 displays a radiosensitizing activity that is highly dependent on treatment modalities by regulating key apoptotic molecular pathways. Our findings therefore support a crucial role for angiogenesis inhibitors in shifting the fate of radiation-induced HIF-1alpha activity from hypoxia-induced tumor radioresistance to hypoxia-induced tumor apoptosis. This study provides a basis for developing new biology-based clinically relevant strategies to improve the efficacy of radiation oncology, using HIF-1 as an ally for cancer therapy.
Figures
References
-
- Schlumberger M. Papillary and follicular thyroid carcinoma. N. Engl. J. Med. 1998;338:297–306. - PubMed
-
- Cooper D.S., et al. Management guidelines for patients with thyroid nodules and differentiated thyroid cancer. Thyroid. 2006;16:109–142. - PubMed
-
- Dai G., Levy O., Carrasco N. Cloning and characterization of the thyroid iodide symporter. Nature. 1996;379:458–460. - PubMed
-
- Eskandari S., et al. Thyroid Na+/I– symporter. Mechanism, stoichiometry, and specificity. J. Biol. Chem. . 1997;272:27230–27238. - PubMed
-
- Filetti S., et al. Sodium/iodide symporter: a key transport system in thyroid cancer cell metabolism. Eur. J. Endocrinol. 1999;141:443–457. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
