

Escherichia coli malic enzymes: two isoforms with substantial differences in kinetic properties, metabolic regulation, and structure
- PMID: 17557829
- PMCID: PMC1952036
- DOI: 10.1128/JB.00428-07
Escherichia coli malic enzymes: two isoforms with substantial differences in kinetic properties, metabolic regulation, and structure
Abstract
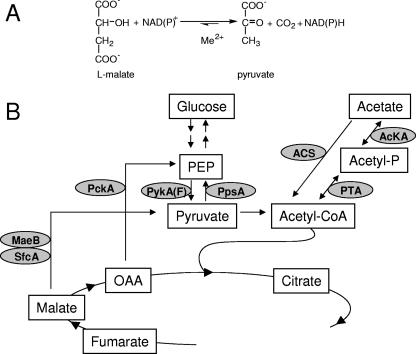
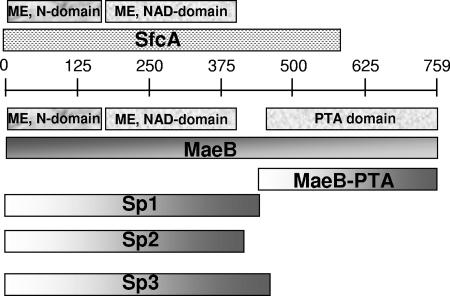
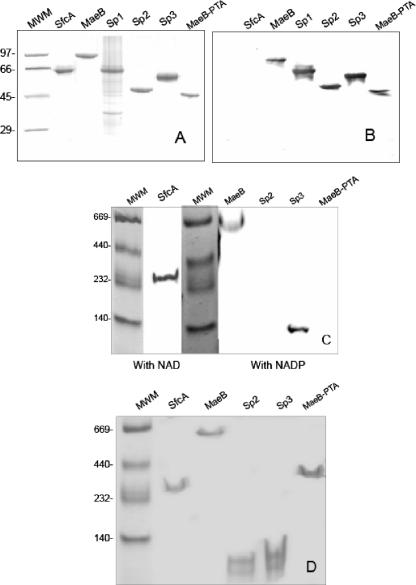
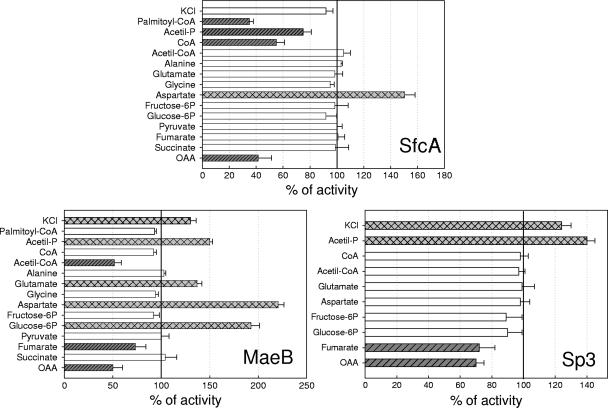
Malic enzymes (MEs) catalyze the oxidative decarboxylation of malate in the presence of a divalent metal ion. In eukaryotes, well-conserved cytoplasmic, mitochondrial, and plastidic MEs have been characterized. On the other hand, distinct groups can be detected among prokaryotic MEs, which are more diverse in structure and less well characterized than their eukaryotic counterparts. In Escherichia coli, two genes with a high degree of homology to ME can be detected: sfcA and maeB. MaeB possesses a multimodular structure: the N-terminal extension shows homology to ME, while the C-terminal extension shows homology to phosphotransacetylases (PTAs). In the present work, a detailed characterization of the products of E. coli sfcA and maeB was performed. The results indicate that the two MEs exhibit relevant kinetic, regulatory, and structural differences. SfcA is a NAD(P) ME, while MaeB is a NADP-specific ME highly regulated by key metabolites. Characterization of truncated versions of MaeB indicated that the PTA domain is not essential for the ME reaction. Nevertheless, truncated MaeB without the PTA domain loses most of its metabolic ME modulation and its native oligomeric state. Thus, the association of the two structural domains in MaeB seems to facilitate metabolic control of the enzyme. Although the PTA domain in MaeB is highly similar to the domains of proteins with PTA activity, MaeB and its PTA domain do not exhibit PTA activity. Determination of the distinct properties of recombinant products of sfcA and maeB performed in the present work will help to clarify the roles of MEs in prokaryotic metabolism.
Figures

References
-
- Brown, D. A., and R. A. Cook. 1981. Role of metal cofactors in enzyme regulation. Differences in the regulatory properties of the Escherichia coli nicotinamide adenine dinucleotide phosphate specific malic enzyme depending on whether Mg2+ or Mn2+ serves as divalent cation. Biochemistry 20:2503-2512. - PubMed
-
- Chang, G.-G., and L. Tong. 2003. Structure and function of malic enzymes, a new class of oxidative decarboxylases. Biochemistry 42:12721-12733. - PubMed
-
- Davis, B. J. 1964. Disc electrophoresis. II. Method and application to human serum proteins. Ann. N. Y. Acad. Sci. 121:404-427. - PubMed
-
- Detarsio, E., M. C. Gerrard Wheeler, V. A. Campos Bermúdez, C. S. Andreo, and M. F. Drincovich. 2003. Maize C4 NADP-malic enzyme: expression in E. coli and characterization of site-directed mutants at the putative nucleotide binding sites. J. Biol. Chem. 278:13757-13764. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
