

Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae
- PMID: 17558514
- PMCID: PMC2084462
- DOI: 10.1007/s00412-007-0111-y
Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae
Abstract
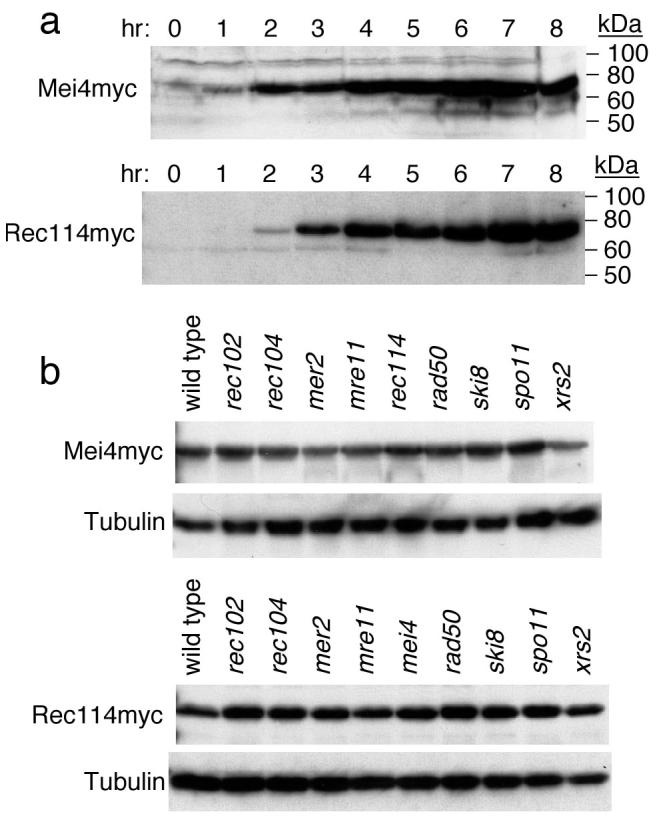
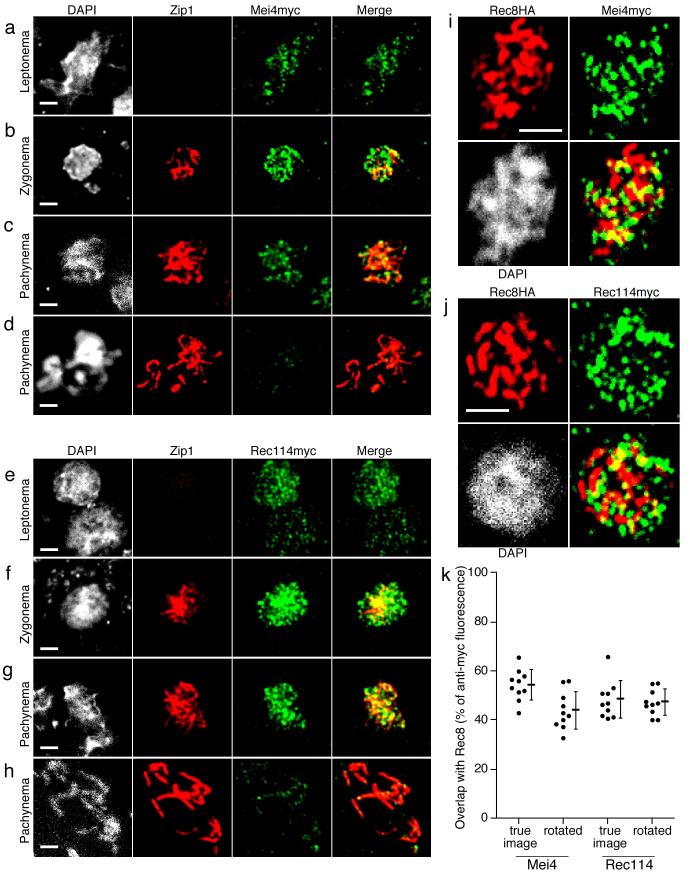
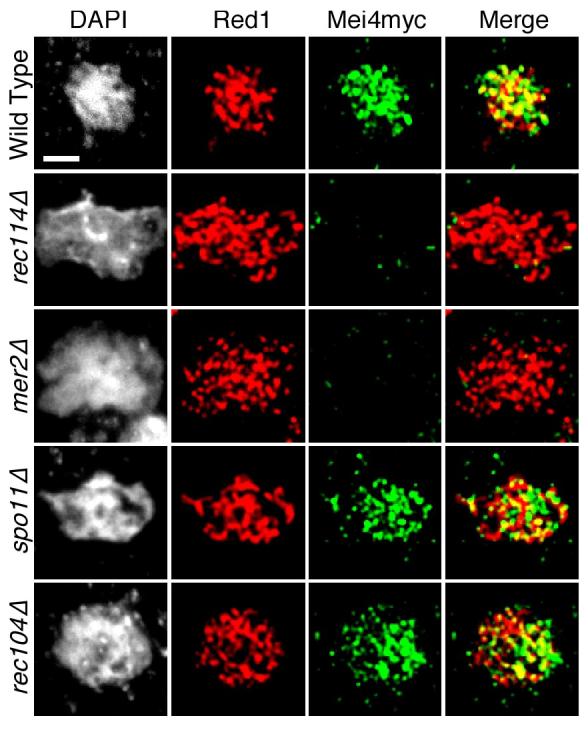
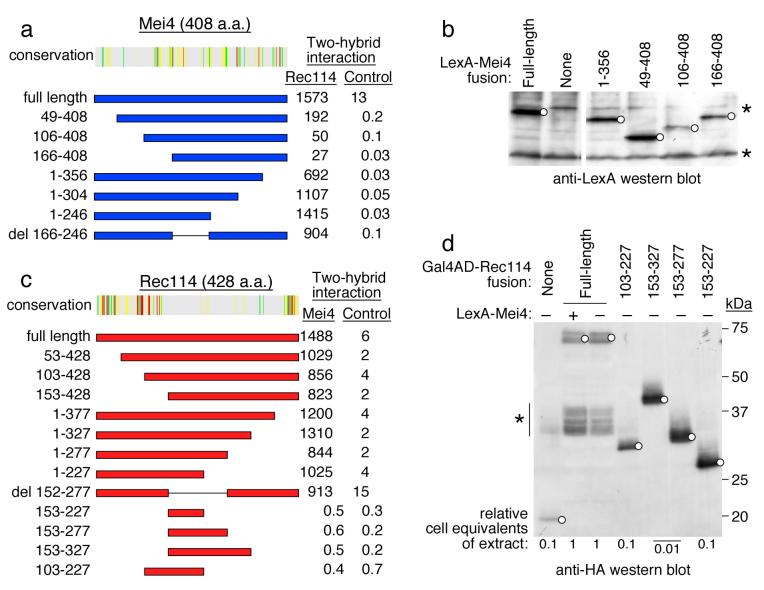
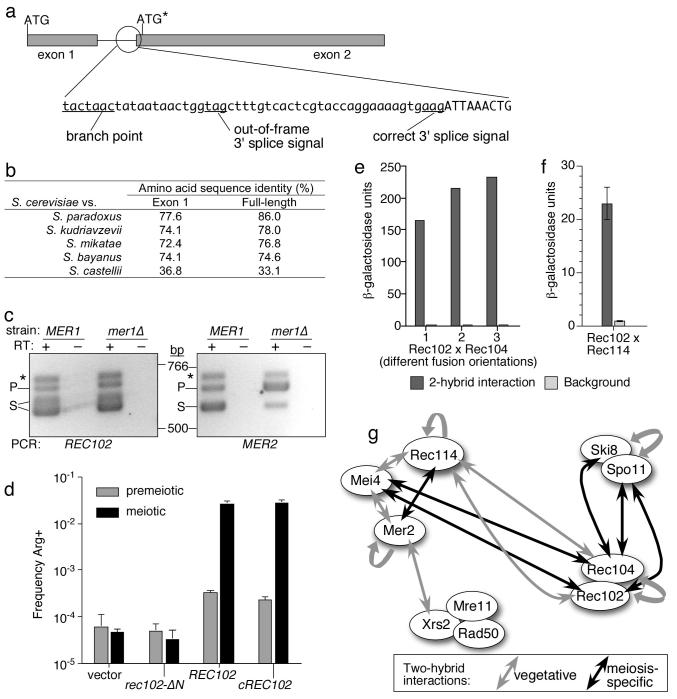
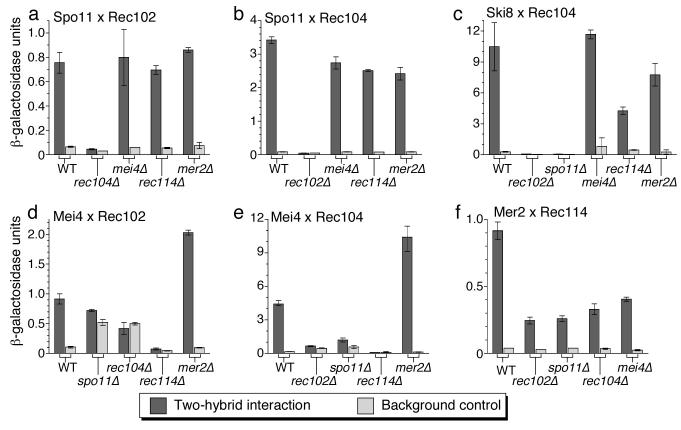
In most sexually reproducing organisms, meiotic recombination is initiated by DNA double-strand breaks (DSBs) formed by the Spo11 protein. In budding yeast, nine other proteins are also required for DSB formation, but the roles of these proteins and the interactions among them are poorly understood. We report further studies of the behaviors of these proteins. Consistent with other studies, we find that Mei4 and Rec114 bind to chromosomes from leptonema through early pachynema. Both proteins showed only limited colocalization with the meiotic cohesin subunit Rec8, suggesting that Mei4 and Rec114 associated preferentially with chromatin loops. Rec114 localization was independent of other DSB factors, but Mei4 localization was strongly dependent on Rec114 and Mer2. Systematic deletion analysis identified protein regions important for a previously described two-hybrid interaction between Mei4 and Rec114. We also report functional characterization of a previously misannotated 5' coding exon of REC102. Sequences encoded in this exon are essential for DSB formation and for Rec102 interaction with Rec104, Spo11, Rec114, and Mei4. Finally, we also examined genetic requirements for a set of previously described two-hybrid interactions that can be detected only when the reporter strain is induced to enter meiosis. This analysis reveals new functional dependencies for interactions among the DSB proteins. Taken together, these studies support the view that Mei4, Rec114, and Mer2 make up a functional subgroup that is distinct from other subgroups of the DSB proteins: Spo11-Ski8, Rec102-Rec104, and Mre11-Rad50-Xrs2. These studies also suggest that an essential function of Rec102 and Rec104 is to connect Mei4 and Rec114 to Spo11.
Figures
References
-
- Alani E, Padmore R, Kleckner N. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell. 1990;61:419–436. - PubMed
-
- Arora C, Kee K, Maleki S, Keeney S. Antiviral protein Ski8 is a direct partner of Spo11 in meiotic DNA break formation, independent of its cytoplasmic role in RNA metabolism. Mol Cell. 2004;13:549–559. - PubMed
-
- Assenmacher N, Hopfner KP. MRE11/RAD50/NBS1: complex activities. Chromosoma. 2004;113:157–166. - PubMed
-
- Blat Y, Protacio RU, Hunter N, Kleckner N. Physical and functional interactions among basic chromosome organizational features govern early steps of meiotic chiasma formation. Cell. 2002;111:791–802. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
