

Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut
- PMID: 17563350
- PMCID: PMC1890564
- DOI: 10.1073/pnas.0704189104
Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut
Abstract
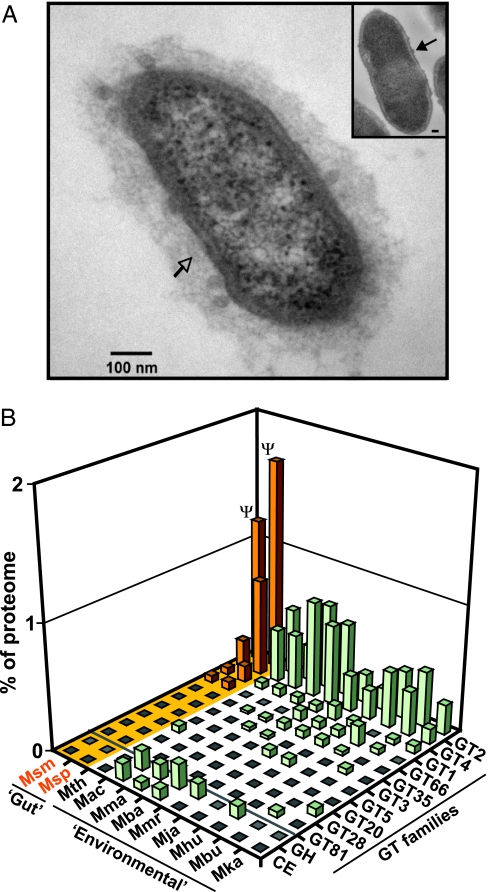
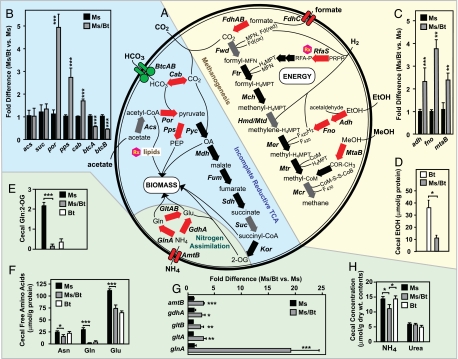
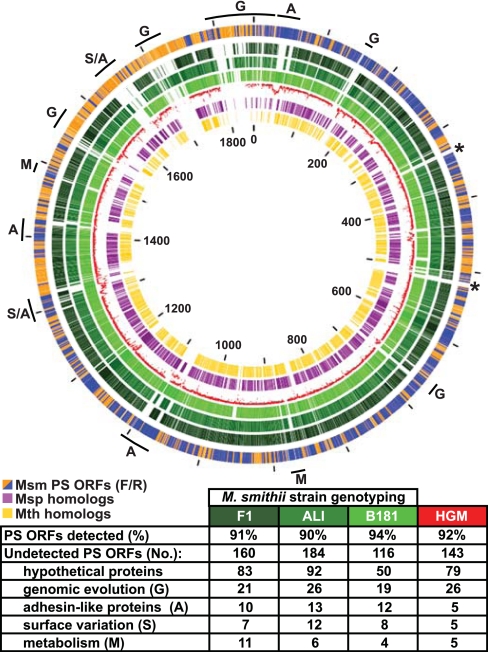
The human gut is home to trillions of microbes, thousands of bacterial phylotypes, as well as hydrogen-consuming methanogenic archaea. Studies in gnotobiotic mice indicate that Methanobrevibacter smithii, the dominant archaeon in the human gut ecosystem, affects the specificity and efficiency of bacterial digestion of dietary polysaccharides, thereby influencing host calorie harvest and adiposity. Metagenomic studies of the gut microbial communities of genetically obese mice and their lean littermates have shown that the former contain an enhanced representation of genes involved in polysaccharide degradation, possess more archaea, and exhibit a greater capacity to promote adiposity when transplanted into germ-free recipients. These findings have led to the hypothesis that M. smithii may be a therapeutic target for reducing energy harvest in obese humans. To explore this possibility, we have sequenced its 1,853,160-bp genome and compared it to other human gut-associated M. smithii strains and other Archaea. We have also examined M. smithii's transcriptome and metabolome in gnotobiotic mice that do or do not harbor Bacteroides thetaiotaomicron, a prominent saccharolytic bacterial member of our gut microbiota. Our results indicate that M. smithii is well equipped to persist in the distal intestine through (i) production of surface glycans resembling those found in the gut mucosa, (ii) regulated expression of adhesin-like proteins, (iii) consumption of a variety of fermentation products produced by saccharolytic bacteria, and (iv) effective competition for nitrogenous nutrient pools. These findings provide a framework for designing strategies to change the representation and/or properties of M. smithii in the human gut microbiota.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
