

Single-stranded oligonucleotide-mediated in vivo gene repair in the rd1 retina
- PMID: 17563719
- PMCID: PMC2765472
Single-stranded oligonucleotide-mediated in vivo gene repair in the rd1 retina
Abstract
Purpose: The aim of this study was to test whether oligonucleotide-targeted gene repair can correct the point mutation in genomic DNA of PDE6b(rd1) (rd1) mouse retinas in vivo.
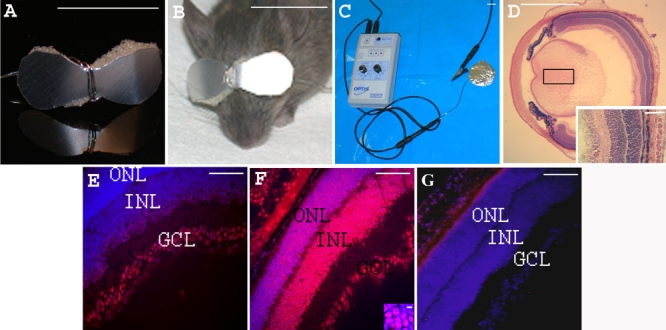
Methods: Oligonucleotides (ODNs) of 25 nucleotide length and complementary to genomic sequence subsuming the rd1 point mutation in the gene encoding the beta-subunit of rod photoreceptor cGMP-phosphodiesterase (beta-PDE), were synthesized with a wild type nucleotide base at the rd1 point mutation position. Control ODNs contained the same nucleotide bases as the wild type ODNs but with varying degrees of sequence mismatch. We previously developed a repeatable and relatively non-invasive technique to enhance ODN delivery to photoreceptor nuclei using transpalpebral iontophoresis prior to intravitreal ODN injection. Three such treatments were performed on C3H/henJ (rd1) mouse pups before postnatal day (PN) 9. Treatment outcomes were evaluated at PN28 or PN33, when retinal degeneration was nearly complete in the untreated rd1 mice. The effect of treatment on photoreceptor survival was evaluated by counting the number of nuclei of photoreceptor cells and by assessing rhodopsin immunohistochemistry on flat-mount retinas and sections. Gene repair in the retina was quantified by allele-specific real time PCR and by detection of beta-PDE-immunoreactive photoreceptors. Confirmatory experiments were conducted using independent rd1 colonies in separate laboratories. These experiments had an additional negative control ODN that contained the rd1 mutant nucleotide base at the rd1 point mutation site such that the sole difference between treatment with wild type and control ODN was the single base at the rd1 point mutation site.
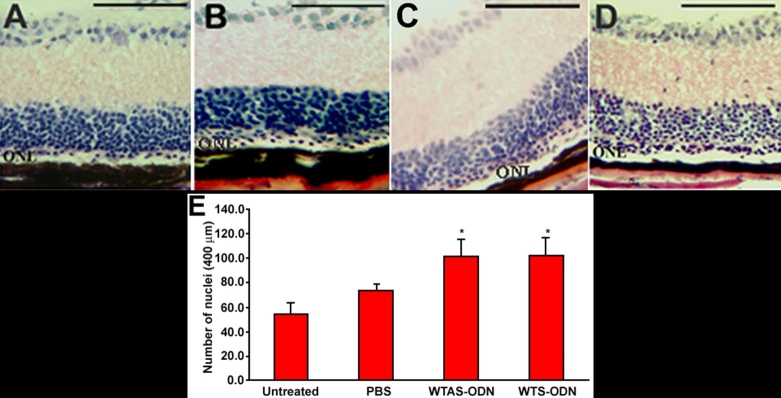
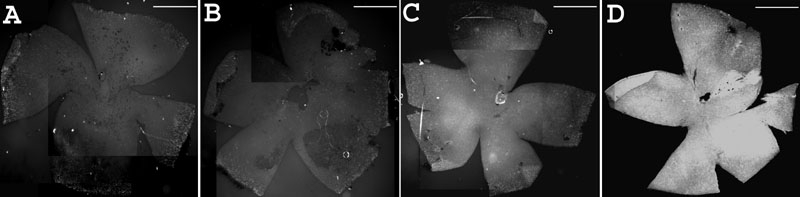
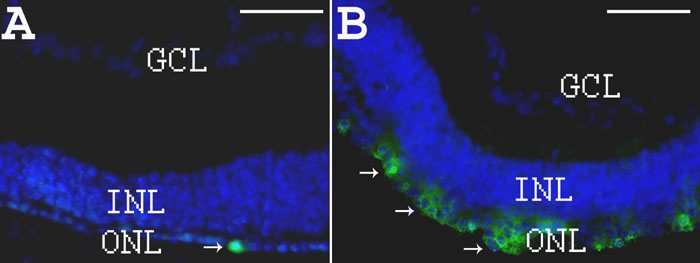
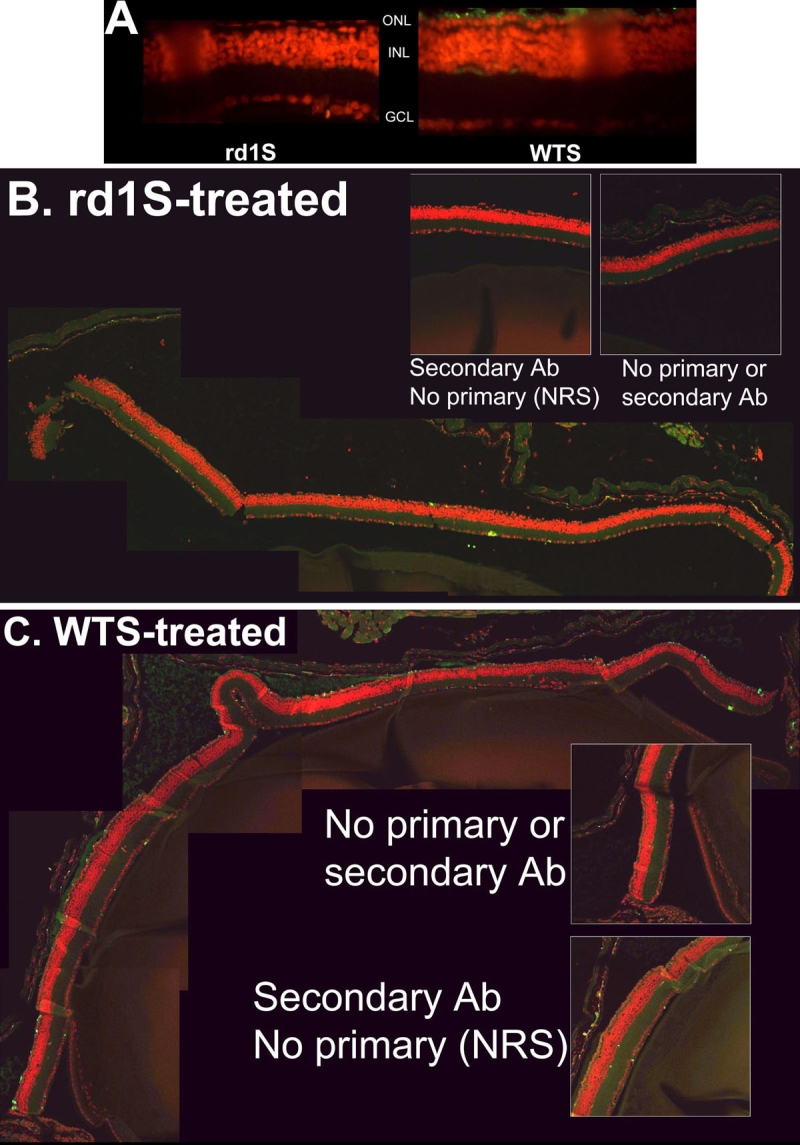
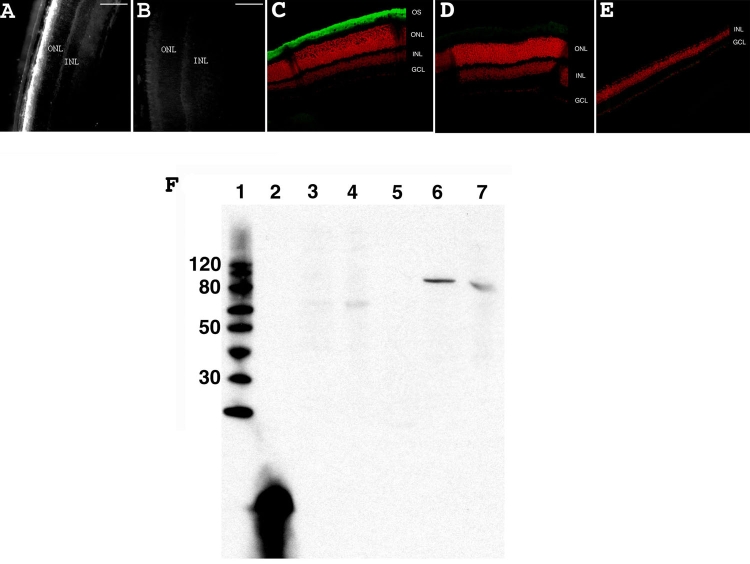
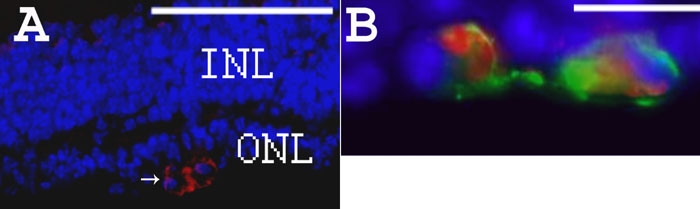
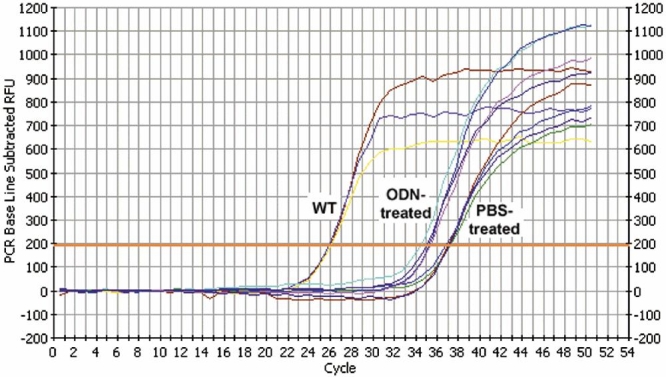
Results: Iontophoresis enhanced the penetration of intravitreally injected ODNs in all retinal layers. Using this delivery technique, significant survival of photoreceptors was observed in retinas from eyes treated with wild type ODNs but not control ODNs as demonstrated by cell counting and rhodopsin immunoreactivity at PN28. Beta-PDE immunoreactivity was present in retinas from eyes treated with wild type ODN but not from those treated with control ODNs. Gene correction demonstrated by allele-specific real time PCR and by counts of beta-PDE-immunoreactive cells was estimated at 0.2%. Independent confirmatory experiments showed that retinas from eyes treated with wild type ODN contained many more rhodopsin immunoreactive cells compared to retinas treated with control (rd1 sequence) ODN, even when harvested at PN33.
Conclusions: Short ODNs can be delivered with repeatable efficiency to mouse photoreceptor cells in vivo using a combination of intravitreal injection and iontophoresis. Delivery of therapeutic ODNs to rd1 mouse eyes resulted in genomic DNA conversion from mutant to wild type sequence, low but observable beta-PDE immunoreactivity, and preservation of rhodopsin immunopositive cells in the outer nuclear layer, suggesting that ODN-directed gene repair occurred and preserved rod photoreceptor cells. Effects were not seen in eyes treated with buffer or with ODNs having the rd1 mutant sequence, a definitive control for this therapeutic approach. Importantly, critical experiments were confirmed in two laboratories by several different researchers using independent mouse colonies and ODN preparations from separate sources. These findings suggest that targeted gene repair can be achieved in the retina following enhanced ODN delivery.
Figures

References
-
- Farber DB, Lolley RN. Cyclic guanosine monophosphate: elevation in degenerating photoreceptor cells of the C3H mouse retina. Science. 1974;186:449–51. - PubMed
-
- Yan W, Lewin A, Hauswirth W. Selective degradation of nonsense beta-phosphodiesterase mRNA in the heterozygous rd mouse. Invest Ophthalmol Vis Sci. 1998;39:2529–36. - PubMed
-
- Farber DB, Danciger M. Identification of genes causing photoreceptor degenerations leading to blindness. Curr Opin Neurobiol. 1997;7:666–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources