

Differentiating between near- and non-cognate codons in Saccharomyces cerevisiae
- PMID: 17565370
- PMCID: PMC1885216
- DOI: 10.1371/journal.pone.0000517
Differentiating between near- and non-cognate codons in Saccharomyces cerevisiae
Abstract
Background: Decoding of mRNAs is performed by aminoacyl tRNAs (aa-tRNAs). This process is highly accurate, however, at low frequencies (10(-3) - 10(-4)) the wrong aa-tRNA can be selected, leading to incorporation of aberrant amino acids. Although our understanding of what constitutes the correct or cognate aa-tRNA:mRNA interaction is well defined, a functional distinction between near-cognate or single mismatched, and unpaired or non-cognate interactions is lacking.
Methodology/principal findings: Misreading of several synonymous codon substitutions at the catalytic site of firefly luciferase was assayed in Saccharomyces cerevisiae. Analysis of the results in the context of current kinetic and biophysical models of aa-tRNA selection suggests that the defining feature of near-cognate aa-tRNAs is their potential to form mini-helical structures with A-site codons, enabling stimulation of GTPase activity of eukaryotic Elongation Factor 1A (eEF1A). Paromomycin specifically stimulated misreading of near-cognate but not of non-cognate aa-tRNAs, providing a functional probe to distinguish between these two classes. Deletion of the accessory elongation factor eEF1Bgamma promoted increased misreading of near-cognate, but hyperaccurate reading of non-cognate codons, suggesting that this factor also has a role in tRNA discrimination. A mutant of eEF1Balpha, the nucleotide exchange factor for eEF1A, promoted a general increase in fidelity, suggesting that the decreased rates of elongation may provide more time for discrimination between aa-tRNAs. A mutant form of ribosomal protein L5 promoted hyperaccurate decoding of both types of codons, even though it is topologically distant from the decoding center.
Conclusions/significance: It is important to distinguish between near-cognate and non-cognate mRNA:tRNA interactions, because such a definition may be important for informing therapeutic strategies for suppressing these two different categories of mutations underlying many human diseases. This study suggests that the defining feature of near-cognate aa-tRNAs is their potential to form mini-helical structures with A-site codons in the ribosomal decoding center. An aminoglycoside and a ribosomal factor can be used to distinguish between near-cognate and non-cognate interactions.
Conflict of interest statement
Figures
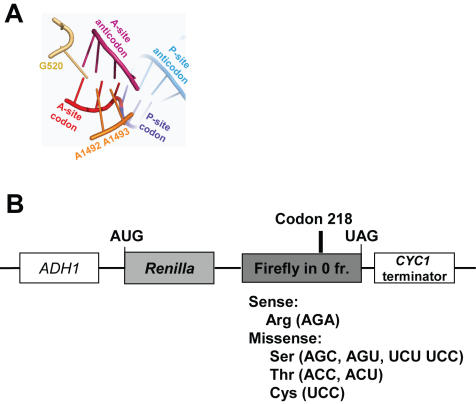
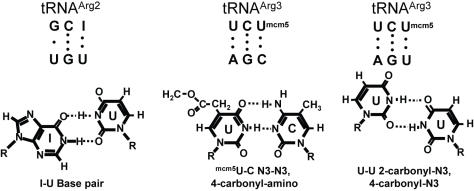
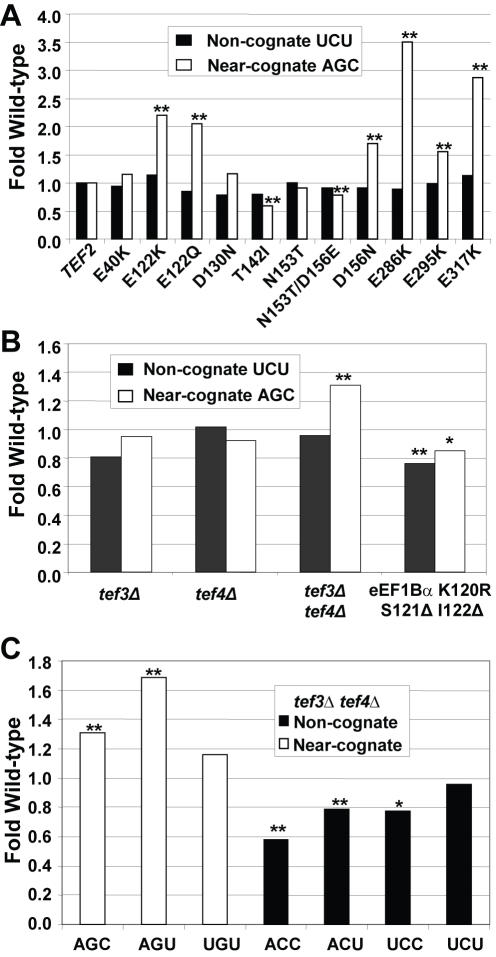
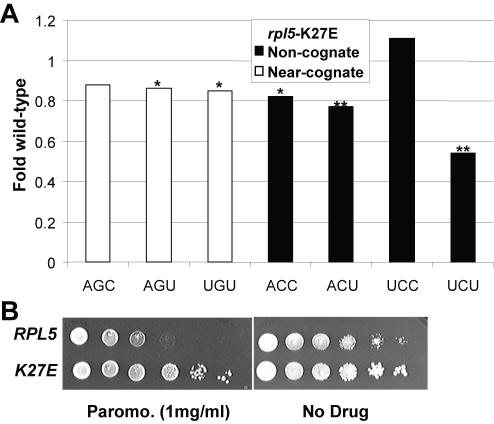
References
-
- Crick FHC. Codon-Anticodon Pairing - Wobble Hypothesis. Journal of Molecular Biology. 1966;19:548–555. - PubMed
-
- Agris PF. Wobble position modified nucleosides evolved to select transfer RNA codon recognition: a modified-wobble hypothesis. Biochimie. 1991;73:1345–1349. - PubMed
-
- Agris PF, Vendeix FA, Graham WD. tRNA's wobble decoding of the genome: 40 years of modification. J Mol Biol. 2007;366:1–13. - PubMed
-
- Ogle JM, Ramakrishnan V. Structural Insights into Translational Fidelity. Annu Rev Biochem. 2005;74:129–177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
