

Cdt1 degradation to prevent DNA re-replication: conserved and non-conserved pathways
- PMID: 17565698
- PMCID: PMC1913051
- DOI: 10.1186/1747-1028-2-18
Cdt1 degradation to prevent DNA re-replication: conserved and non-conserved pathways
Abstract
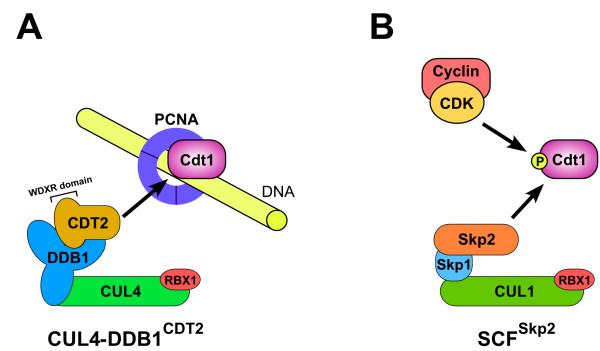
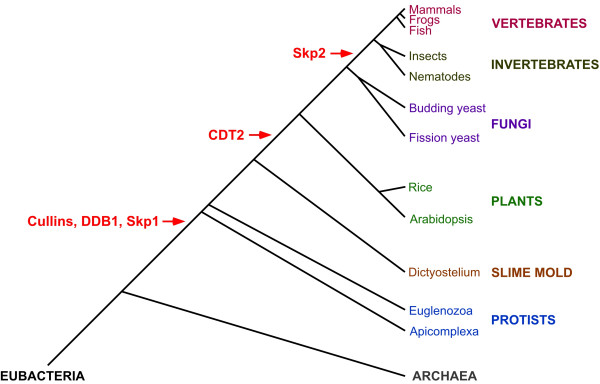
In eukaryotes, DNA replication is strictly regulated so that it occurs only once per cell cycle. The mechanisms that prevent excessive DNA replication are focused on preventing replication origins from being reused within the same cell cycle. This regulation involves the temporal separation of the formation of the pre-replicative complex (pre-RC) from the initiation of DNA replication. The replication licensing factors Cdt1 and Cdc6 recruit the presumptive replicative helicase, the Mcm2-7 complex, to replication origins in late M or G1 phase to form pre-RCs. In fission yeast and metazoa, the Cdt1 licensing factor is degraded at the start of S phase by ubiquitin-mediated proteolysis to prevent the reassembly of pre-RCs. In humans, two E3 complexes, CUL4-DDB1CDT2 and SCFSkp2, are redundantly required for Cdt1 degradation. The two E3 complexes use distinct mechanisms to target Cdt1 ubiquitination. Current data suggests that CUL4-DDB1CDT2-mediated degradation of Cdt1 is S-phase specific, while SCFSkp2-mediated Cdt1 degradation occurs throughout the cell cycle. The degradation of Cdt1 by the CUL4-DDB1CDT2 E3 complex is an evolutionarily ancient pathway that is active in fungi and metazoa. In contrast, SCFSkp2-mediated Cdt1 degradation appears to have arisen relatively recently. A role for Skp2 in Cdt1 degradation has only been demonstrated in humans, and the pathway is not conserved in yeast, invertebrates, or even among other vertebrates.
Figures
References
-
- Feng H, Kipreos ET. Preventing DNA re-replication – divergent safeguards in yeast and metazoa. Cell Cycle. 2003;2:431–434. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
