

Nuclear interactions of topoisomerase II alpha and beta with phospholipid scramblase 1
- PMID: 17567603
- PMCID: PMC1919507
- DOI: 10.1093/nar/gkm434
Nuclear interactions of topoisomerase II alpha and beta with phospholipid scramblase 1
Abstract
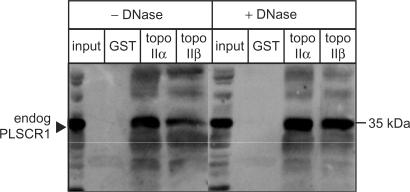
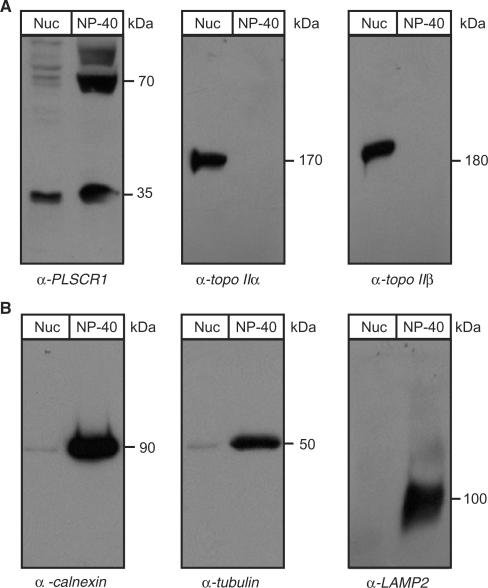
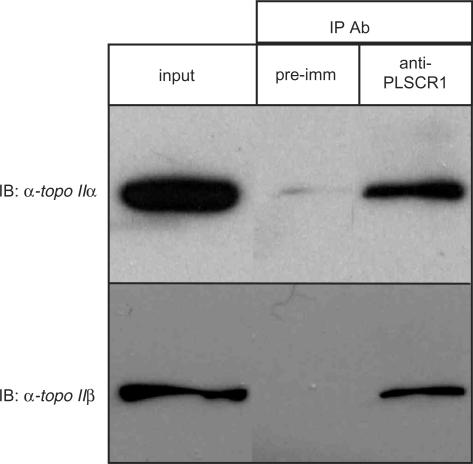
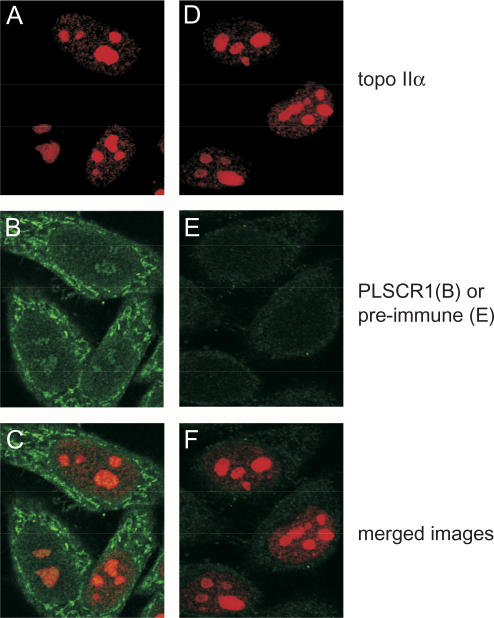
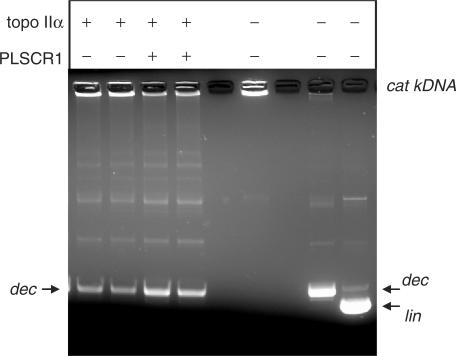
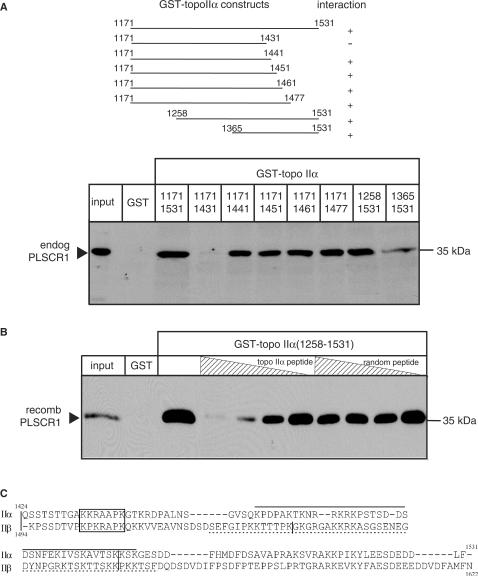
DNA topoisomerase (topo) II modulates DNA topology and is essential for cell division. There are two isoforms of topo II (alpha and beta) that have limited functional redundancy, although their catalytic mechanisms appear the same. Using their COOH-terminal domains (CTDs) in yeast two-hybrid analysis, we have identified phospholipid scramblase 1 (PLSCR1) as a binding partner of both topo II alpha and beta. Although predominantly a plasma membrane protein involved in phosphatidylserine externalization, PLSCR1 can also be imported into the nucleus where it may have a tumour suppressor function. The interactions of PLSCR1 and topo II were confirmed by pull-down assays with topo II alpha and beta CTD fusion proteins and endogenous PLSCR1, and by co-immunoprecipitation of endogenous PLSCR1 and topo II alpha and beta from HeLa cell nuclear extracts. PLSCR1 also increased the decatenation activity of human topo IIalpha. A conserved basic sequence in the CTD of topo IIalpha was identified as being essential for binding to PLSCR1 and binding of the two proteins could be inhibited by a synthetic peptide corresponding to topo IIalpha amino acids 1430-1441. These studies reveal for the first time a physical and functional interaction between topo II and PLSCR1.
Figures
References
-
- Wang JC. Cellular roles of DNA topoisomerases: a molecular perspective. Nat. Rev. Mol. Cell Biol. 2002;3:430–440. - PubMed
-
- Woessner RD, Mattern MR, Mirabelli CK, Johnson RK, Drake FH. Proliferation- and cell cycle-dependent differences in expression of the 170 kilodalton and 180 kilodalton forms of topoisomerase II in NIH-3T3 cells. Cell Growth Differ. 1991;2:209–214. - PubMed
-
- Bauman ME, Holden JA, Brown KA, Harker WG, Perkins SL. Differential immunohistochemical staining for DNA topoisomerase II α and β in human tissues and for DNA topoisomerase IIβ in non-Hodgkin's lymphomas. Mod. Pathol. 1997;10:168–175. - PubMed
-
- Watanabe M, Tsutsui K, Inoue Y. Differential expressions of the topoisomerase IIα and IIβ mRNAs in developing rat brain. Neurosci. Res. 1994;19:51–57. - PubMed
-
- Tsutsui K, Sano K, Kikuchi A, Tokunaga A. Involvement of DNA topoisomerase IIβ in neuronal differentiation. J. Biol. Chem. 2001;276:5769–5778. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
